Обмен веществ (метаболизм) дрожжей
Микроорганизмы осуществляют свои жизненные функции, главным проявлением которых является рост и размножение за счет многочисленных непрерывно происходящих и взаимосвязанных химических реакций, обычно называемых обменом веществ или метаболизмом. Метаболизм представляет собой два типа процессов, а именно:
анаболизм или ассимиляция характеризуется синтетическими реакциями, при которых потребляется энергия, организмы принимают из среды вещества (питание), эти вещества они способны переработать в продукты, компенсирующие потери, вызванные потреблением и расщеплением, и пригодные для образования новых веществ для роста и размножения;
катаболизм или диссимиляция, т. е. сумма реакций разложения, богатых энергией высокомолекулярных веществ, на вещества простейшие, бедные энергией, а освобождающаяся энергия потребляется на процессе ассимиляции.
Метаболизм сахаридов
В обмене веществ дрожжей и всех живых систем вообще большое значение имеют сахара (сахариды), при расщеплении которых получается энергия для физиологических процессов и для биологического синтеза многих веществ. Расщепление сахаров (гликолиз) является одним из основных процессов. Если он протекает при прямом участии кислорода воздуха, т. е. при аэробных условиях, таких, как окислительный обмен веществ, речь идет о процессе, обычно называемом дыханием (respirace). При анаэробных условиях брожение (ферментация) протекает как антиокислительный обмен веществ.
Дыхание и брожение - это ферментативные процессы, сложный механизм которых в начальных стадиях подобен. Они протекают вначале через ряд промежуточных стадий, а затем на определенной стадии разделяются и идут к конечным продуктам своими собственными путями. При дыхании конечным продуктом являются углекислый газ и вода
C6H12O6+6O2=6CO2+6H2O+2830 кДж (674 ккал)
и при брожении спирт (этанол) и углекислый газ
C6H12O6->2C2H5OH+2CO2+924 кДж (22 ккал).
Количество энергии, освободившейся при этих катаболических (диссимиляционных) процессах, зависит от степени окисления. Чем выше теплота сгорания образующихся продуктов, тем меньше энергии освободилось для клетки. Метаболизм сахаридов (углеводов) и его отношение ко всему обмену веществ выражает следующая схема [5]:
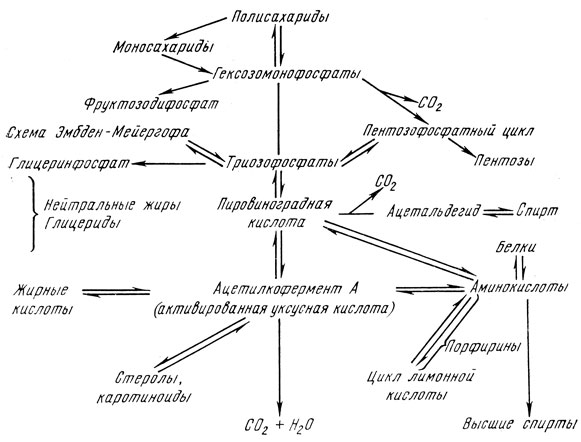
Полисахариды и олигосахариды (мальтоза, сахароза) или сначала расщепляются на моносахариды и фосфорилируются, прямо переходят в фосфаты гексозы. Последний из них фруктозо-1,6-дифосфат расщепляется на два фосфорилированных С3-сахара (по схеме Эмбден-Мейергофа) или уже на предшествующей стадии глюкозо-6-фосфат дегидрогенизируется на 6-фосфоглюконовую кислоту и путем окислительной декарбоксилации перейдет в пентозофосфат, который является голозидным компонентом рибонуклеиновых кислот. Фосфаты пентозы в так называемом пентозофосфатном цикле переходят через гептозы, гексозы, тетрозы в фосфаты триоз и гексоз, через которые присоединяются к схеме Эмбден-Мейергофа.
Из триозофосфата (дигидроксиацетонфосфата) образуется в качестве побочного продукта глицерин, основное вещество для синтеза нейтральных жиров и фосфатидов.
Основной путь расщепления сахаридов ведет от триозофосфата через разные промежуточные стадии к пировиноградной кислоте. При декарбоксилации и восстановлении образуются основные продукты анаэробного гликолиза, т. е. этанол и углекислый газ. Кроме того, пировиноградная кислота, отдает свой углеродный скелет аминокислоте аланина, который образуется в результате трансаминирования.
При аэробных условиях протекает также процесс окисления, в результате которого образуется "активированная уксусная кислота". При скоплении ацетиловых остатков активированной уксусной кислоты образуются жирные кислоты, стеролы, каротиноиды и одновременно образуется углеродный скелет ряда важных аминокислот. Эти процессы обратимы и поэтому ацетилкофермент А является важным промежуточным веществом расщепления, перестройки и построения сахаридов, белков, жиров, стеролов, каротиноидов и других веществ.
Окончательное превращение ацетиленового остатка в углекислый газ и воду осуществляется в цикле лимонной кислоты, благодаря чему клетка приобретает основную часть энергии. Из промежуточных продуктов α-кетоглютаровой и щавелевоуксусной кислот при аминировании и трансаминировании образуются глютаминовая и аспарагиновая кислоты.
При пивоваренном брожении основным и преобладающим процессом является анаэробный гликолиз (брожение) по схеме Эмбден-Мейергофа-Парнасова, при котором в качестве основных продуктов образуются этанол и углекислый газ. В присутствии кислорода на процесс брожения накладываются аэробные процессы (дыхание), в частности в стадии пировиноградной кислоты цикл лимонной кислоты (Кребса) или в стадии глюкозо-6-фосфата-пентозофосфатный цикл.
При низовом пивоваренном брожении приблизительно 98% сахара сбраживается и 2% используются на процесс дыхания.
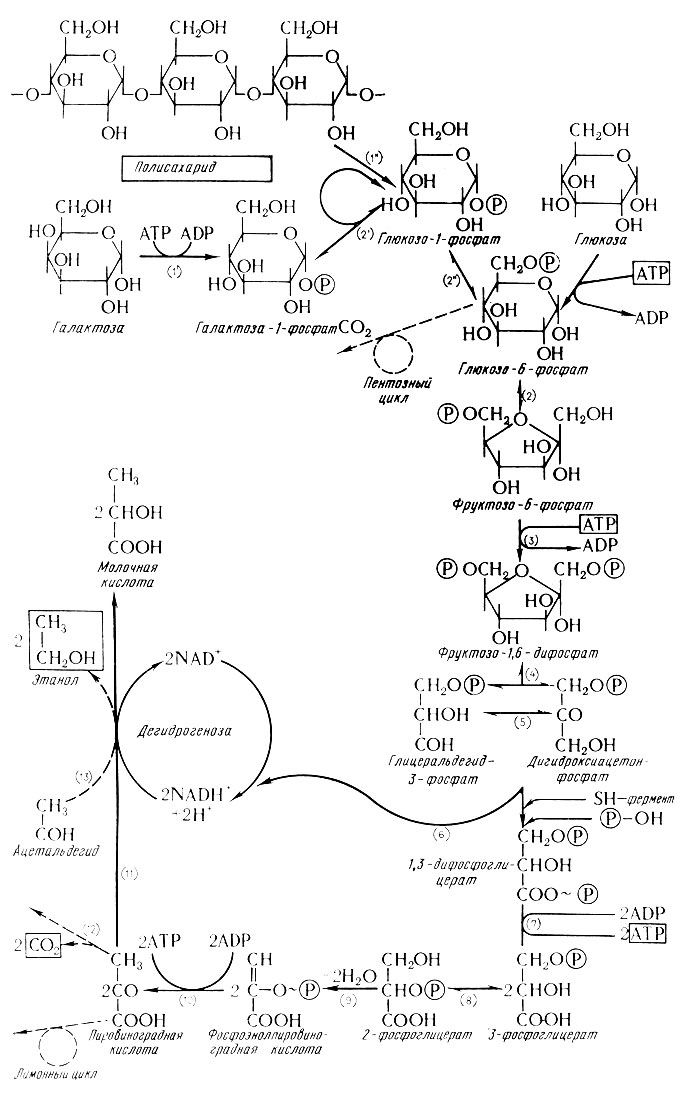
Спиртовое брожение (схема Эмбден-Ме йергофа)
Спиртовое брожение очень сложный процесс, при изучении которого постепенно выяснилось, что он протекает как цепь ре-акций, связанных фосфорилирующим механизмом.
В первой фазе брожения (стадия фосфорилирования) образуются разные фосфаты гексоз. Последний из них (фруктозо-1,6- дифосфат) распадается на два триозофосфата: дигидроксиацетонфосфат и глицеральдегид-3-фосфат. Глицеральдегид-3-фосфат дегидрогенизируется при вмешательстве никотинамидадениндинуклеотида (НАД) и одновременно фосфорилируется в 1,3-фосфорглицериновую кислоту, которая последующей дегидрогенизацией и дефосфорилизацией переходит в пировиноградную кислоту. Кислота в свою очередь декарбоксилируется в углекислый газ и ацетальдегид, который в последней фазе восстанавливается в спирт (этанол). Весь ход спиртового брожения виден по при-веденной по Де Клерку (6) схеме, которая составлена на основе имеющихся данных, относящихся к изучению этого сложного процесса.
Сбраживаемость сахаров различна и зависит от вида сахара и от свойств дрожжей. Сначала считали, что прямо сбраживаются только так называемые зимогексозы, т. е. глюкоза, фруктоза, маноза и условно (после приспособления) также галактоза, при этом все только в оптической D-форме. Олигосахариды считались сбраживаемыми только после расщепления на гексозы соответствующими ферментами. Например, низовые пивоваренные дрожжи содержат инвертазу (сахарозу, β-фруктозидазу-β-D-фруктофуранозид фруктогидролазу, ЕК 3.2.1.26), катализирующую расщепление сахарозы на глюкозу и фруктозу, далее мальтозу (α-глюкозидазу-α-D-глюкозид глюкогидролазу, ЕК 3.2.1.21), которая катализирует расщепление мальтозы на две молекулы глюкозы и, наконец, мелибиазу (α-галактозидазу-α-D-галактозид галактогидролазу, ЕК 3. 2. 1. 23), которая при одновременном присутствии сахарозы расщепляет трисахарид на все три его составляющие, т. е. галактозу, глюкозу и фруктозу. У верховых дрожжей мелибиаза отсутствует и поэтому они расщепляют рафинозу на мелибиозу и фруктозу, так что сбраживают только ее фруктозный компонент (1/3).
В последнее время с помощью хроматографического метода подтвердилось существование трансфераз, способных переносить гексозные единицы с одной молекулы дисахарида или трисахарида на другую, или также внутри молекулы с одной связи на другую. Так, под действием трансглюкозидазы может образоваться например, из сахарозы трисахарид фруктозо-фруктозо-глюкоза или фруктозо-глюкозо-фруктоза. Образовавшиеся таким образом сахара под действием трансфераз расщепляются (сбраживаются) непосредственно ферментами брожения. Сбраживаемость сахаров зависит также от проницаемости клеточной оболочки дрожжей. Известно, что внутрь дрожжей проникают дитрисахариды, а высшие сахара и декстрины никогда.
Согласно суммарному уравнению брожения из 100 г глюкозы теоретически получается 51,1 г спирта (этанола) и 48,9 г углекислого газа. Практически выход всегда ниже и соответствует в среднем 48,6% этанола и 46,4% углекислого газа. Около 2% глюкозы дрожжи ассимилируют и остаток приблизительно 3% переходит в побочные продукты, т. е. ацетилдегид, глицерин, уксусную и молочную кислоту.
Цикл лимонной кислоты
В начале брожения сусло всегда содержит определенное количество растворенного кислорода воздуха, в присутствии которого дрожжи "дышат". Тем самым изменяется первоначальная схема анаэробного метаболизма. Восстановление ацетилдегида замедляется, так что начинается скапливаться пировиноградная кислота, незначительный излишек которой аэробно декарбонизируется в цикле лимонной кислоты, или в трикарбоксиловом цикле, называемом также по имени его открывателя циклом Кребса.
В реакцию включается следующий комплекс ферментов, катализирующий в присутствии кофермента А образование ацетил- кофермента А из очень небольшого количества пировиноградной кислоты. При его действии на щавелевоуксусную кислоту образуется лимонная кислота, а из нее постепенно кислоты цисаконитовая, L-изолимонная, щавелевоянтарная, α-кетоглутаровая, фумаровая, L-яблочная и, наконец, щавелевоуксусная, которая остается в цикле. Ее реакцией с пировиноградной кислотой начинается новый цикл.
Уже раньше было известно, что при полном исключении дыхания снижается жизнеспособность пивных дрожжей, а тем самым снижаются и их физиологические функции и бродильная активность. Вайнфуртнер [3] приводит, как вывод из новых исследований [15], сведения о том, что энергия, получаемая при дыхании, физиологически неравноценна энергии, полученной при брожении, поскольку без дыхания дрожжи постепенно утрачивают способность к размножению.
Пентозофосфатный цикл
В последнее время обнаружилось, что при дыхании дрожжей до 30% глюкозы расщепляется прямым окислением [16] в следующем так называемом пентозофосфатном или гексозомонофосфатном цикле по схеме Варбурга, Дикенса и Хорекера. При спиртовом брожении он накладывается на схему Эмбден-Майергофа в стадии глюкозо-6-фосфата, который окисляется в 6-фосфоглюконовую кислоту (через δ-лактон). При декарбоксилации через 6-фосфо-3-кетоглутаровую кислоту образуется рибулозо-5- фосфат. Последняя образует с рибозо-5-фосфатом седогептулозо-7-фосфат и глицеральдегид-3-фосфат, который возвращается в нормальную схему Эмбден-Мейергофа. Седогептулозо-7-фосфат расщепляется в эритрозо-4-фосфат и дигидроксиацетон. В присутствии глицеральдегид-3-фосфата образуется фруктозо-6-фосфат, который начинает следующий цикл.
Одним кольцом пентозофосфатного цикла одна молекула глюкозо-6-фосфата декарбонизуется в рибулозо-5-фосфат. При этом восстанавливаются две молекулы трифосфопиридиннуклеотида (ТПН), который снова должен окисляться кислородом воздуха.
Эффект Пастера
Основные изменения в метаболизме дрожжей, связанном с подачей воздуха в процессе брожения (анаэробный и аэробный процесс), первым заметил Пастер. Он обнаружил, что кислород подавляет брожение (образование спирта), однако ускоряет рост (размножение) дрожжей. Это явление известно как эффект Пастера. При аэробных условиях дрожжи не сбраживают (кроме пивных) сахар в этанол, а образуется только углекислый газ; при этом по сравнению с аэробным гликолизом потребление глюкозы намного меньше. По теории Линена [17], диапазон расщепления сахара регулирует концентрация неорганического фосфата и аденозиндифосфата (АДФ).
У пивоваренных культурных дрожжей эффект Пастера не наблюдается [18]. С технологической точки зрения наличие эффекта Пастера свидетельствовало бы о нарушении функций пивоваренных дрожжей.
Метаболизм азотистых веществ
Азотистые вещества в дрожжевых клетках постоянно обновляются. Одновременно с расщеплением протекает синтез белков. В качестве азотистых продуктов питания дрожжевым клеткам служат соли аммония, аминокислоты и низшие пептиды, если они проникают через оболочку клетки.
Для роста, обновления и восстановления живых систем особое значение имеют аминокислоты. Они образуются при введении аминогруппы в α-кетокислоты, которые являются промежуточными продуктами при расщеплении сахаридов. Преимущественную часть потребления аминокислот (около 75%) дрожжи осуществляют прямой ассимиляцией и почти весь остаток дезаминированием по схеме Эрлиха с образованием высших спиртов [19]. Для синтеза белков наиболее важна глютаминовая и аспарагиновая кислоты, аланин и амиды первых двух, т. е. глютамин и аспарагин.
Кроме аминокислот дрожжи ассимилируют также пептиды [20]. Из азота, ассимилированного из сусла дрожжами, около 11 % приходится на пептиды. Поэтому значение пептидов по сравнению с соответствующими аминокислотами меньше, а именно потому, что под действием экзопептидаз они сначала должны расщепляться в простые компоненты.
Внутри дрожжевых клеток аминокислоты подвергаются трансаминированию катализованному трансаминозами, переносящими аминогруппы с одной молекулы на другую. Так, например, аланин образуется путем реакции глютаминовой кислоты с пировиноградной. Высокомолекулярные азотистые вещества образуются конденсацией аминокислот при участии аденозинтрифосфата (АТФ). В пивном сусле аминокислоты являются более важным источником азота, чем аммониевые соединения [21]. Легко ассимилируемыми являются аргинин, лизин, метионин, глютаминовая и аспарагиновая кислоты. И, наоборот, пролин, валин и другие выделяются в среду.
Сусло всегда содержит достаточное количество азотистых веществ. Из 600 мг ассимилированного азота, содержащегося в 1 л сусла (12%-ное), при брожении дрожжи потребляют около 200 мг [22]. В стадии роста содержание азота в дрожжах быстро возрастает, пока не достигнет почти двукратной величины [23] и потом снова постепенно падает вплоть до исходной величины. Дальнейшее уменьшение содержания азота связано с тем, что дрожжи выделяют в среду от 5 до 33% из собственного объема азота, вероятнее всего в виде аминокислот и пептидов.
Метаболизм минеральных веществ
Общее значение минеральных веществ в клеточном организме вытекает из следующих функций:
они поставляют основные (неорганические) элементы для построения органических соединений (углерод, азот, сера и фосфор);
регулируют и поддерживают в организме определенные величины некоторых физических или физико-химических свойств, например прочность клеточной оболочки, осмотическое давление протоплазмы, ионное равновесие, pH, проницаемость клеточной оболочки, растворимость белков, активность ферментов и т. д.;
являются составной частью некоторых биологических катализаторов, главным образом ферментов (железо, медь), витаминов (кобальт), красителей (магний), активируют или ингибируют многие ферментативные реакции (магний, железо, медь, кальций, калий, натрий, кобальт, цинк и т. д.) и тем самым прямо вмешиваются в метаболические процессы.
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://wine.historic.ru/ 'Виноделие как искусство'