Часть 3. ВИРУСНЫЕ И ВИРУСОПОДОБНЫЕ БОЛЕЗНИ МАЛИНЫ И ЕЖЕВИКИ
ВВЕДЕНИЕ
РАСПРОСТРАНЕНИЕ И ОБНАРУЖЕНИЕ ВИРУСНЫХ БОЛЕЗНЕЙ РАСТЕНИЙ РОДА RUBUS И БОРЬБА С НИМИ
Культивируемые виды Rubus подразделяются на две большие группы: малину и ежевику. Их часто называют ягодными кустарниками.
Малина относится к подроду Idaeobatus, зрелые ягоды ее легко отделяются от цветолож, на которых они образуются. Обычно возделываются два вида малины: красная, или обыкновенная, малина (Rubus idaeus L.) и черная, или западная, малина (R. occidenta-lis L.). Американские фо'рмы красной малины часто помещают в вид R. idaeus L. var. strigosus (Michx.) Maxim. Некоторые специалисты гибриды между красной и черной малиной называют пурпуровой малиной и относят ее к отдельному виду R. neglectus Peck. Часть форм пурпуровой малины выращивают в промышленном масштабе. Черную малину обычно возделывают во многих частях США. Красная малина выращивается в отдельных районах северной и южной умеренных зон, в частности в Европе, Северной Америке, Австралии и Новой Зеландии.
Ежевика относится к подроду Eubatus. Спелая ягода, состоящая из цветолож и сросшихся мелких костянок, отделяется как целое с плодоножкой. Имеется много видов ежевики и межвидовых гибридов, которые выращивают в виде сортов. Трудно и иногда невозможно перечислить дикие виды, которые участвовали в происхождении многих распространенных сортов.
Согласно характеру роста ежевика подразделяется на две большие группы: прямостоячую и стелющуюся ежевику, или росянику. Обе часто называются сложноплодными. Некоторые формы стелющейся ежевики, например логанова ягода и Бойзенберрп, произошли от красной малины и ежевики или росяники. Ежевику возделывают в тех же районах, как и малину, но ее ареал включает районы, несколько более близкие к субтропикам, чем у малины.
Большей частью эти ягодные растения являются основной культурой лишь в отдельных областях, например в Восточной Шотландии и на северо-западе США. Ежегодное производство ягод культивируемых видов малины и ежевики в Европе и Северной Америке оценивается Министерством сельского хозяйства США в 33 млн. долл.
Свыше 20 вирусных болезней, которые поражают культивируемые и дикие виды Rubus, описывается в некоторых деталях в этом справочнике. Другие вирусные и вирусоподобные заболевания были описаны ранее, но еще недостаточно изучены для детального рассмотрения. Некоторые из этих второстепенных вирусов и вирусоподобных болезней кратко рассматриваются в отдельной статье этой части.
Не все вирусные болезни растений Rubus вызывают серьезные потери урожая. Одни наносят повреждения растениям, но не имеют значения из-за ограниченного географического распространения. Другие не опасны вследствие толерантности их хозяев. Однако некоторые вирусные болезни растений Rubus очень сильно распространены и наносят большой ущерб. Таким образом, вирусные болезни представляют собой главную проблему, которая ограничивает возделывание этих ягодных культур.
Превосходные обзоры о вирусах растений Rubus были опубликованы Стейс-Смитом [223] и Кадменом [45]. Дискуссия о вирусных болезнях также включена в сельскохозяйственный справочник №310 о болезнях растений Rubus, опубликованный Министерством сельского хозяйства США [69].
Группы вирусов растений Rubus. В данном справочнике вирусы растений Rubus сгруппированы согласно их естественным способам передачи. Это сделано для удобства пользования справочником и не обязательно отражает какие-либо взаимоотношения. Физические и химические свойства ряда вирусов, переносимых нематодами, известны. В противоположность этому, способы естественного распространения вирусов, вызывающих такие важные заболевания, как полосатость черной малины, стерильность и карликовость ежевики, не известны. Такие вирусные заболевания объединены в громоздкую и разнородную группу и произвольно помещены в одном месте.
Изучение вирусов растений Rubus, вероятно, будет продолжаться в течение ряда лет большей частью в полевых, чем в лабораторных условиях. Однако большие достижения могут быть получены от изучения самих вирусов, а также от применения методов современной вирусологии и серологии.
Мнение, что большинство вирусов древесных растений имеет ограниченный круг хозяев, преобладало в прошлом у вирусологов по плодовым культурам. Для растений Rubus была показана ошибочность такого представления на примере появления таких вирусов, как кольцевой пятнистости томата, мозаики резухи и хлороти-ческой пятнистости листьев яблони на различных сортах Rubus. По этой причине приемы и индикаторы, которые пригодны для изучения вирусов, поражающих другие растения семейства розоцветных, должны использоваться для изучения вирусов, заражающих растения Rubus.
Меры борьбы. Устойчивые сорта в конечном счете обеспечивают лучшую борьбу с вирусными болезнями Rubus. Одним из препятствий к более полному использованию генов устойчивости к этим вирусам является отсутствие точности в их идентификации. Пока вирусологи и генетики по Rubus не достигнут уровня точности, необходимого для выведения устойчивых сортов, до тех пор получение клонов, свободных от определенных опасных вирусов, размножение и распространение этих клонов садоводами по хорошо разработанным программам сертификации, по-видимому, будет наиболее надежным средством для снижения потерь урожая.
Холлингс [130] сделал превосходный обзор по производству свободного от вирусов клопового посадочного материала. Отбор и термотерапия использовались для получения высококачественного посадочного материала Rubus, свободного от многих вирусов.
Чамберс [61] первым применил термотерапию для освобождения посадочного материала малины от некоторых латентных вирусов. Он освободил ряд английских сортов красной малины от термолабильных компонентов вируса мозаики (ТЛКМ) путем выращивания растений в освещаемых камерах при 35°С [62]. Болтон и Турнер [20] применили термотерапию с последующим размножением малины верхушками побегов для получения растений, свободных от мозаики. Конверс [67, 69] использовал подобные приемы для черной, красной и пурпуровой малины в США. Такого рода работы выполнены во Франции [177], в Нидерландах и в Скандинавских странах. Некоторые сорта красной малины освобождены от вируса карликовости Rubus методом термотерапии [244].
Не все вирусы, поражающие Rubus, являются термолабильными. Некоторые ценные сорта полностью заражены термостабильными вирусами, такими, как термостабильные компоненты мозаики (ТСКМ), и латентным вирусом черной малины. Культура меристем и хемотерапия являются экспериментальными методами, доступными в дополнение к термотерапии для освобождения растений от вирусов. В некоторых случаях при работе с другими культурами все три метода или комбинации из них успешно применялись для освобождения клонов от вирусов. Такой комбинированный подход только начинает использоваться в терапии вирусов Rubus и является многообещающим.
Программа сертификации видов Rubus основана на использовании растений, полученных путем индексирования, отбора и термотерапии, и они, насколько возможно, свободны от вирусов. Клоны таких линий размножаются в условиях, при которых возможность повторной инфекции сведена до минимума. Эта система является динамичной, поскольку более совершенными методами иногда обнаруживают в маточном материале новые вирусы. Улучшенные методы терапии вирусов позволяют получать новый здоровый маточный материал, доступный садоводам. Исходным растениям, выпускаемым Шотландским научно-исследовательским институтом садоводства и Министерством сельского хозяйства США, присваиваются кодовые номера, которые облегчают идентификацию сертифицированных клонов данного сорта и обеспечивают внедрение клонов более высокого качества [4, 67].
Хотя термин "свободный от вирусов" применяется часто без разбора к выделенным, отобранным или индексированным клонам, но даже, примененный к лучшим клонам, он является абсолютным статичным термином, который не способствует успешному исследованию вируса данной культуры. Неопределенный термин "проверенный на вирус", предпочитаемый в английских свидетельствах сертификации, более приемлем [79].
Поскольку мозаика малины является наиболее важной вирусной болезнью в роде Rubus, против которой направлена программа сертификации, можно оправдать использование термина "свободный от мозаики". Однако в настоящее время этот термин часто используется в промышленных питомниках для обозначения клонов малины, которые не проявляют симптомов мозаики, хотя они могут быть носителями латентных вирусов мозаики. Применение термина "свободный от мозаики" для обозначения бессимптомно зараженных мозаикой растений малины должно обескураживать потребителя.
Поддержание материнских растений для программы сертификации Rubus лежит на специальных агентствах. Такие клоны передают квалифицированным питомниководам для размножения с соблюдением минимально принятой изоляции и мер борьбы с вредителями. В практике существует несколько уровней сертификации посадочного материала, соответствующих увеличивающемуся числу вегетационных периодов, в течение которых он размножается с момента получения из блока маточных растений. Хорошими примерами схем сертификации Rubus служат схемы, которые применяются в Шотландии [5], в провинции Онтарио [6] и штате Нью-Йорк (США) [23].
Поддержание правильной изоляции, хорошая борьба с наземными и почвенными переносчиками и удаление цветков для предотвращения распространения вируса с пыльцой - важные аспекты правильного руководства питомником.
Для рентабельного промышленного выращивания малины садоводу необходимо применять сертифицированный посадочный материал и учитывать два обстоятельства: 1) скорость распространения вируса в местных условиях должна быть медленной или контролируемой, или же ущерб, причиняемый урожаю, должен быть минимальным; 2) стоимость и доступность исходного или размноженного посадочного материала должны быть такими, чтобы садовод мог вести дело с прибылью.
Если эти два условия обеспечены, садоводы могут быть заинтересованы в обновлении посадок сравнительно часто и таким образом избегать накопления вирусных болезней. Вирусы будут накапливаться, если одна и та же плантация сохраняется в течение многих лет.
Приемы индексирования. Опыты по передаче инфекции прививкой, тлями и механическим путем на восприимчивые к вирусу индикаторные растения - три способа, используемые для определения присутствия латентных вирусов в клонах Rubus.
Опыты по передаче прививкой. Харрис [НО] был первым исследователем, продемонстрировавшим возможность переноса мозаики прививкой в расщеп, пластинкой коры и сближением. Кадмен [49] использовал модификацию бутылочного способа прививки побега сближением и в некоторых случаях получил симптомы на индикаторных растениях через две недели после прививки. Ван дер Меер [173] показал, что вирус крапчатости листьев (ТЛКМ)* мог быть с успехом передан клинообразной прививкой черешков от листочков больного растения в расщеп черешка листочка на индикаторном растении. Конверс [68] применил способ прививки листом для передачи ТЛКМ и ТСКМ 1 на Rubus henryi Hemsl. et Kuntze. Для обнаружения ТЛКМ (и, вероятно, ТСКМ) при 5%-ном уровне достоверности необходимо, чтобы три привитых листочка на тест-растение оставались жизнеспособными в течение 14 дней. За индикаторным растением велось наблюдение еще в течение 21 дня, и если симптомы не развивались, то растение-донор считалось свободным от мозаики. Как растение-донор, так и индикаторное растение должны быть мощными по развитию, чтобы добиться передачи вирусов мозаики.
*(ТЛКМ - термолабильный компонент вируса мозаики. ТСКМ - термостабильный компонент вируса мозаики. - Ред.)
Опыты по передаче тлями. Передача вируса с помощью тли Amphorophora agathonica Hottes, обычно относимой к A. rubi (Kalt.), была использована Стейс-Смитом [219, 220] в качестве метода индексации для обнаружения вирусов ТЛКМ и ТСКМ (см. стр. 176). Очень небольшие, мощные сеянцы черно.й малины были использованы в качестве тест-растений. Хубер [139] описал метод сбора в полевых условиях образцов побегов с подозрительных на заболевание растений малины для индексирования с помощью тлей. Конверс [70] установил, что предельная эффективность групп тлей A. agathonica в переносе мозаики малины на черную малину равна 76% по сравнению с 97% при прививке листочком на Rubus henryi. Применение нескольких сеянцев черной малины достаточно для индексирования каждого растения (относительно простая процедура с тлями); индексирование мозаики малины с помощью тлей также является быстрым и надежным методом. Известны другие тли-переносчики вирусов Rubus, но они не использовались для обычных целей индексирования.
Опыты по механической передаче. Обнаружение вирусов хлоротичной пятнистости листьев яблони, кольцевой пятнистости томата, европейских вирусов Rubus, сохраняющихся в почве, и латентного вируса черной малины на красной малине наиболее удобно провести путем механической передачи соком с подозрительного растения на травянистые растения-индикаторы. Для успешной передачи вируса соком с зараженных растений Rubus существует по меньшей мере три важных фактора: 1) использование сока из листьев молодых хорошо растущих побегов, предпочтительно нового прироста, или весной из частей цветков; 2) инокуляция должна быть проведена на сочных, мощных травянистых тест-растениях, таких как Chenopodium quinoa Willd; 3) тест-растения следует выращивать в прохладной (16 - 24°С) и влажной теплице, частично затеняемой летом. Часто в северных районах с умеренным климатом тест-растения, выращиваемые в теплице зимой, являются наиболее подходящими. Если любой из этих трех факторов не находится в оптимуме, то могут быть только следы инфекции или она совсем не будет обнаружена даже при заведомо зараженных растениях Rubus.
Добавление 2%-ного никотина в воду со свежеотжатым соком Rubus помогает поддержать щелочную реакцию его и предотвращает естественную инактивацию вирусов, особенно при содержании танинов в соке [41]. Согласно Кадмену, алюминий действует подобно никотину, но менее эффективен.
Такие приемы, как добавление абразивов (часто применяются целит, диатомовая земля) и сильное затенение индикаторных растений на несколько часов после инокуляции, увеличивают процент передачи при работе с вирусами Rubus.
При механической передаче вирусов малины должны соблюдаться некоторые меры предосторожности. 1. Семена R. occidentalis и, вероятно, других видов Rubus обычно несут один из механически переносимых вирусов: хлоротичную пятнистость листьев яблони и латентный вирус черной малины. Эти вирусы являются латентными и термостабильными на R. occidentalis, но вызывают системный некроз или хлоротичную крапчатостъ и кольцевую пятнистость на С. quinod, [Конверс и Кадмен; не опубликовано; 71].
2. Несколько видов Chenopodium, включая С. quinoa, иногда несут переносимый семенами вирус мозаики С. rubrum [14]. Этот вирус обычно латентен до тех пор, пока хозяин не поврежден, например абразивом при механической передаче [8]. Вирус мозаики С. rubrum, называемый также вирусом звездчатой крапчатости Chenopodium, имеет изометрические частицы, накапливается в высокой концентрации, термостабилен, переносится механически [8,151].
3. Вирус табачной мозаики часто встречается, очевидно, как случайный загрязнитель во время проведения опытов с вирусами растений в теплице. Он найден в деревьях яблони и груши в естественных условиях [105, 156].
В опытах по механической передаче вирусов, поражающих Rubus, необходимо доказывать отсутствие любого из этих вирусов как загрязнителей.
Тест-растения. Хотя некоторые вирусы образуют характерные симптомы на своих растениях-хозяевах рода Rubus, многие находятся либо в латентном состоянии, либо выявляют только кратковременные или слабозаметные симптомы. Тест-растения, перечисленные в таблице 6, были использованы несколькими исследователями в качестве индикаторов латентных вирусов, но еще не разработан систематический или диагностический список индикаторов. Использование этих хозяев и симптомы, обнаруживаемые на них, описаны подробно при обсуждении отдельных болезней в этой части.

Таблица 6. Некоторые важные растения-индикаторы для определения вирусов на растениях Rubus
МОЗАИКА МАЛИНЫ (RASPBERRY MOSAIC)
Определение. Мозаика малины рассматривается как категория заболеваний, включающая те вирусные болезни Rubus, которые: а) переносятся между растениями-хозяевами Rubus тлями Amphorophora rubi (Kalt.) и A. agathonica Hottes* и б) вызывают шоковые симптомы, состоящие из некротизации верхушек побегов и (или) крапчатости, если переносятся на чувствительные клоны Rubus henryi Hemsl. et Kuntze или R. occidentalis L. Вирусы хлороза жилок [32] и желтухи [34] не рассматриваются как часть группы вирусов мозаики малины, поскольку они не передаются тлями Amphorophora.
* (Amphorophora agathonica Hottes - вид тлей, к которому Л. Руссель (персональное сообщение) отнесены все коллекции североамериканских тлей, прежде определенные как A. rubi (Kalt.). Поскольку размещение этих видов тлей является предварительным или временным, A. agathonica в дальнейшем в этой части справочника будет указываться как A. agathonica (?).)
Синонимы. Большое разнообразие симптомов на различных растениях-хозяевах Rubus, вызываемых вирусами, более или менее соответствующих приведенному выше определению, приводит к необычно большому числу названий болезни. Многие из этих названий являются местными или относятся к заболеваниям, для которых мало известно о вирусах возбудителях.
Так как источники многих болезней, описанных в литературе, отсутствуют, невозможно отнести их к любой из трех больших категорий групп мозаики малины, перечисленных ниже. Поэтому они разбиты на четыре категории. Мы рассматриваем мозаику малины, включающую расстройства, вызываемые: 1) термолабильными компонентами мозаики; 2) термостабильными компонентами мозаики; 3) комплексной инфекцией, в которую вовлечены термолабильный и термостабильный компоненты, давая начало обыкновенной мозаике малины; 4) изолятами неизвестной термостабильности.
1. Термолабильные компоненты мозаики (ТЛКМ) [66] Некроз черной малины (НЧМ) [220] Крапчатость листьев малины [31] Пятнистость листьев малины [35] Хлоротичная пятнистость листьев малины [150] 2. Термостабильные компоненты мозаики (ТСК.М) Желтая мозаика [11] Желтая сетчатость Rubus (ВЖСК) [219] 3. Обыкновенная мозаика малины Это болезнетворный комплекс, обусловленный присутствием обоих (термолабильного и термостабильного) вирусов мозаики. Мозаика малины = вирус некроза черной малины плюс вирус желтой сетчатости Rubus [221], или = ТЛКМ плюс ТСКМ. Окаймление жилок [33] = мозаика малины в понимании Стейс-Смита [45]. 4. Свойства термостабильности не известны Зелено-крапчатая мозаика = зеленая мозаика [76]) Мозаика малины I [110] Мозаика малины II (слабая) [НО] Мозаика малины II (сильная) [НО] Пятнистая мозаика [193] Окаймление жилок [198] Тринадцать различных типов мозаики, основанных на симптомах [125]
Вопросы о взаимоотношениях среди многих перечисленных мозаичных заболеваний и действительном числе вовлеченных вирусов могут быть решены лишь после дальнейшего определения свойств отдельных вирусов.
История и географическое распространение. В Европе, где малина культивируется с середины XVI в., на дикой малине, удаленной от культивируемых сортов Rubus, часто обнаруживают мозаику. Это же относится к восточной части Северной Америки, где малина стала возделываться только 200 лет назад.
С 1900 по 1920 г. в Северной Америке мозаику и курчавость листьев смешивали между собой, и часто относили к одной болезни "желтухе". Мозаика была четко отделена от курчавости листьев в 1920-х годах. Ранние статьи о мозаике малины были опубликованы Беннетом [11], Харрисом [108, 110] и Стейс-Смитом [221]. Уилкокс и Смит [255] были первыми, кто доказал, что Amphorophora agathonica (?) является переносчиком мозаики. Aphis rubicola Oestlund (= rubiphila Patch) также рассматривалась некоторыми учеными как переносчик. В классической статье Беннетт [11] показал, что A. rubicola не служит переносчиком вируса мозаики малины.
Мозаика малины в широком смысле встречается везде на выращиваемых сортах Rubus, особенно на красной, черной и пурпуровой малине. Симптомы не везде одни и те же. Часто этим заболеваниям даются специфические названия, поскольку они встречаются в данном районе в зависимости от выращиваемых сортов и имеющихся штаммов вируса. Мозаика малины, очевидно, не так широко распространена в Калифорнии, Орегоне и Вашингтоне, как в остальных штатах США.
Экономическое значение. Сообщений о влиянии мозаики малины или ее компонентов на рост и урожай малины очень мало [66]. Урожай ягод у двух сортов красной малины на востоке США снижался под воздействием ТЛКМ на 53% по сравнению с клонами, свободными от мозаики [66]. Подобные данные по красной малине из Шотландии [262] показывают, что по крайней мере вначале отсутствие вирусов мозаики сказывается на усилении энергии роста и повышении потенциальной урожайности. Нет точных данных о влиянии мозаики на рост и урожайность черной или пурпуровой малины и ежевики. Однако, по общим наблюдениям, вегетативный рост, урожай и качество ягод черной малины ухудшаются под влиянием мозаичной инфекции, тогда как вред пурпуровой малине и ежевике незначителен.
Круг растений-хозяев
Естественные растения-хозяева Rubus, subgenus Idaeobatus R. idaeus L., широко распространена R. innominatus S. Moore [254] R. leucodermis Dougl. [140, 265] R. occidentalis L. и гибриды с красной малиной, широко распространены R. odoratus L. [265] R. parviflprus Nutt. [265] R. phoenicolasius Maxim., обычный дикий хозяин, локально [63, 265] R. spectabilis Pursh [265] R. idaeus L. var strigosus (Michx.) Maxim., широко распространена Rubus, subgenus Eubatus R. laciniatus Willd. [265, Кадмен; не опубликовано] R. loganobaccus Bailey [265] R. macropetalus Dougl. [265] R. procerus P. J. Muell. [219, Кадмен, не опубликовано] Rubus species Различные названные и не имеющие названия, культивируемые и дикие, прямостоячие и стелющиеся виды ежевики [11, Кадмен, не опубликовано]. Нет сообщений о том, чтобы естественными хозяевами были растения вне рода Rubus. Экспериментальные хозяева Семейство Rosaceae при прививке или с участием переносчика Rubus albescens Roxb. [219] R. henryi Hemsl. et Kuntze [31, Чамберс; не опубликовано] R. ellipticus Smith [Конверс; не опубликовано] R. saxatilis L. [31] Fragaria vesca L. (только для ВЖСН) [229] Травянистые растения-хозяева при передаче соком (только ТЛКМ) [11, 110, 125, 248] Chenopodium amaranticolor Coste et Reyn [44] С. quinoa Willd. [44, 198] Datura stramonium L. [44] Petunia hybrida Vilm. [44] Vigna sinensis (Torner) Savi, использованный Фултоном (102) в качестве экспериментального хозяина для мозаики малины, был, вероятно, инфицирован вместо мозаики вирусом кольцевой пятнистости томата.
До тех пор пока исследователи смогут переносить вирус с травянистых тест-растений обратно на малину и вызывать характерные симптомы мозаики, эти сообщения должны рассматриваться только как обнадеживающие, но предварительные данные о новых травянистых хозяевах вирусов мозаики малины. Всегда следует иметь в виду, что имеются латентные, переносимые семенами вирусы Rubus, которые легко передаются соком на Chenopodium quinoa, но не переносятся тлей Amphorophora agathonica (?) [71, 74].
Симптомы. Неблагоприятные погодные условия, особенно поздние весенние заморозки, иногда способствуют образованию крапчатости на листьях плодоносящих побегов, которая очень похожа на мозаику [11]. Однако листья молодых побегов, развившиеся в конце весны, обычно выглядят нормальными. Мучнистая роса также может иммитировать симптомы мозаики на листьях, но гриб-возбудитель образует белый мучнистый налет и водянистые участки на нижней стороне листа. На симптомы мозаики малины могут также походить повреждения листьев, вызванные инвазией красного клеща, или тлей Amphorophora rubitoxica Knowlton [217], или опрыскиванием определенными фунгицидами, или симптомы недостаточности бора в почве [170].
Разнообразие симптомов очень характерно для мозаики, даже на одном сорте Rubus. Географическое местоположение, условия вегетационного сезона и особенности сорта влияют на проявление симптомов. Имеется обширная литература о варьировании симптомов мозаики малины, но мало критических сведений о их этиологии. Какими вирусами вызываются мозаичные болезни Rubus, покажут будущие исследования, когда будут разработаны улучшенные методы идентификации вирусов. Симптомы, вызываемые некоторыми изолятами вируса мозаики, уже описывались в Великобритании Харрисом [108, ПО], Прентисом и Харрисом [191], Кадменом [31, 33, 35] и в Северной Америке Беннеттом [И], Ранкином [195, 196], Шумейкером и др. [209], Стейс-Смиттом [219, 220, 221].
Ниже приведено описание симптомов мозаики малины, встречающейся в Северной Америке.
Термолабильные компоненты мозаики (ТЛКМ).Черная малина, вероятно, является культурой, которая наиболее сильно поражается как термолабильными, так и термостабильными компонентами вируса мозаики. Первоначальный некроз верхушечных листьев и верхушек побегов (рис. 63, вверху), вызываемый ТЛКМ, сопровождается крапчатостью нижних листочков (рис. 63, внизу). Хронически инфицированные растения могут развивать мелкие, крапчатые листья на карликовых побегах. Новые побеги могут образовывать розетку. Степень проявления первоначальной некротической реакции варьирует от незаметной до интенсивной [70]. Крапчатость также может варьировать от слабой до сильной; низкие температуры способствуют проявлению симптомов [И].
Красная малина. Симптомы неотчетливые. Как начальные, так и хронические симптомы варьируют от слабых хлоротических пятен вдоль боковых жилок до полного их отсутствия. Неясная крап-чатость может развиваться на новом, весеннем приросте. Многие из основных сортов красной малины в Северной Америке сплошь инфицированы [64, 220]; такое же положение, вероятно, наблюдается и в Европе. Энергия роста у зараженных растений снижается, основное влияние заключается в уменьшении числа побегов на единицу площади. Густота стояния растений сорта Ньюбург, пораженного ТЛКМ, составила 49% от таковой здоровых растений [66].

Рис. 63. Вверху - симптомы начальной инфекции на побеге сорта Мюнгер, пораженного вирусом некроза черной малины, который является термолабильным компонентом вируса мозаики. Внизу - хроническая крапчатость листа, развивающаяся после вышеуказанных симптомов
Ежевика. Немного работ проведено по изучению вирусов мозаики на ежевике. Изредка наблюдалась резкая крапчатость. Установлено, что некоторые растения, зараженные ТЛКМ, были бессимптомными, как определено индексацией на Rubus henryi и тепловой обработкой [219, Конвере; не опубликовано].
Термостабильные компоненты мозаики (ТСКМ). Эта группа вирусов, по крайней мере на североамериканских сортах красной малины, в естественных условиях почти всегда встречается вместе с ТЛКМ [66, 221].
Поэтому описываемые здесь симптомы выявлены при экспериментальном заражении.
Черная малина. Первоначальные симптомы представляют собой расплывчатые хлоротичные пятна вдоль жилок на субтерминальных листочках. Эти пятна сливаются с хлорозом жилок, образуя сплошной, обычно асимметричный, сетевидный узор, который приводит к некоторой деформации листьев (рис. 64, слева) [221]. "Желтая мозаика" может быть выражением хронических симптомов ТСКМ на черной малине, но анализ симптомов и изучение термотерапии отсутствуют. Симптомы желтой мозаики в полевых условиях описаны Беннеттом [11]: больные растения карликовые и слабые; листья отчетливо желтые, и края их могут закручиваться вниз, но на листьях не проявляются крапчатость и пузырчатые участки. Симптомы сохраняются в жаркую погоду. Ягоды на таких растениях семенистые, крошащиеся и негодные для употребления. Зараженные растения живут недолго.
Красная малина. Шоковые симптомы не развиваются. Хронические симптомы выражены бледным, сетевидным хлорозом жилок и слабой чашевидностью листочков (рис. 64, в центре). Болезнь мало отражается на росте растений и их урожае [221].

Рис. 64. Слева и в центре - листья черной малины и красной малины сорта Вашингтон, зараженные вирусом желтой сетчатости малины, термостабильным компонентом вируса мозаики. Справа - симптомы мозаики малины (ТЛКМ+ТСКМ) на листе красной малины сорта Кутберт
Ежевика. Желтая сетчатость: первый вирус из группы ТСКМ был выделен и описан [219] с инфицированных в естественных условиях растений ежевики сорт Гималайя (Rubus procerus P. J. Muell.), на которых он вызывал ярко-желтое посветление жилок, часто развивающееся асимметрично на листьях, и чашевидность листочков. Вегетативный рост и урожайность заметно не снижались. Термостабильные компоненты мозаики были обнаружены только однажды в питомнике ежевики в 1966 г. на растениеводческой станции в Белтсвилле, штат Мэриленд (Конверс; не опубликовано).
Обыкновенная мозаика малины.Черная малина. Весной зараженные растения покрываются листвой на несколько дней позднее, чем здоровые растения [78]. Верхушки побегов часто некротические и ломкие. Побеги короткие .и расположены розетками, хотя рост может быть нормальным во время жаркой погоды. Ягоды мелкие и семенистые. Растения обычно отмирают в течение нескольких лет.
Красная малина. Листья, образовавшиеся в холодную погоду, крапчатые, пузырчатые и искривленные (рис. 64, справа). Листья, развившиеся в жаркую погоду, могут быть бессимптомными. Вегетативный рост снижается, а урожай и качество ягод ухудшаются [221].
Ежевика. См. ТЛКМ. Сильнокрапчатые листья иногда встречаются на культивируемых и диких растениях ежевики как в США, так и в Европе, но анализ симптомов не проводили.
Характеристика вируса. Не было технических описаний вирусов, известных в качестве возбудителей мозаики малины. Кадмен [45] сообщил о неокончательных результатах опытов по перекрестной защите между термолабильными компонентами мозаики, хотя ранее он полагал [33], что только опыты подобного рода могут установить взаимоотношения между вирусами малины. Стейс-Смит [218] решил, что два штамма вируса мозаики (вероятно, ТЛКМ) не полностью защищают один от другого, но присутствие слабого штамма вызывало задержку развития симптомов от сильного штамма, если им последовательно заражали того же хозяина; это указывает на степень интерференции и, следовательно, на определенную взаимосвязь.
Передача. Мозаика малины экспериментально передается прививкой, возможно механической инокуляцией сока и тлями. Эти вирусы не передаются ни через почву, ни семенами.
а) Передача прививкой. Передача мозаики была осуществлена прививками в расщеп, пластинкой коры, сближением [110], бутылочным способом [31] илистом [173].
Конверс [68] показал, что, если три прививки листочком с растений малины, инфицированных ТЛКМ или ТСКМ, остаются жизнеспособными в течение 14 дней на Rubus henryi, вирусы могут быть обнаружены на индикаторе с достоверностью 95%.
Типично, что ТЛКМ вызывает некроз верхушки у инокулированного прививкой R. henryi и инокулированного тлей R. occidentalis в течение 7 - 10 дней, в то время как ТСКМ вызывает посветление жилок, крапчатость и деформацию листьев у инокулированного прививкой R. henryi в течение 20 дней, за которыми следует некроз верхушки побегов и почек [68]. Однако некоторые изоляты ТСКМ вызывают начальный некроз листочков на R. henryi. Один необычный изолят, предположительно отнесенный к ТСКМ, вызвал сильный и быстро развившийся некроз верхушки побега R. henryi после трех последовательных тепловых обработок растения-хозяина (сорт красной малины Латам), но не мог быть передан на R. occidentalis с помощью тлей [70].
б) Механическая передача инфекционным соком. Ни один вирус из группы ТСКМ еще не был передан соком. Вирусы из группы ТЛКМ, вероятно, с успехом передавались механически с малины на Chenopodium quinoa и некоторые другие травянистые растения-хозяева Кадменом [42, 44] и Рихтером [198, 200]. Обратная инокуляция ТЛКМ на Rubus с травянистых растений и даже серия пассажей на травянистые растения не была достигнута. Бернайер и Кинг [17] сообщили о передаче двух механически переносимых вирусов малины тлей Amphorophora agathonica (?). Последующее обнаружение в одной из этих культур вируса табачной мозаики (Бернайер, персональное сообщение) делает ранние заключения авторов сомнительными.
в) Передача с помощью переносчика. Взаимосвязь переносчика и вируса мозаики малины была описана в обзоре Кеннеди и др. [195], и только некоторые тли были определены как переносчики.
Вирусы мозаики малины являются полуперсистентными в этих переносчиках [232]. Взаимоотношения вирус - переносчик для двух видов тлей рассмотрены в таблице 7.

Таблица 7. Взаимоотношения между изолятами мозаики малины и двумя видами тлей-переносчиков
Amphorophora rubi (Kalt.) subsp. idaei Born - главный переносчик вирусов мозаики малины в Европе [51, 155]. Macrosiphum euphorbiae (Thos.), возможно, имеет в Европе меньшее значение [36]. Свенсон и Милбрат [230] рассматривали эту тлю как вероятного переносчика мозаики малины на побережье Тихого океана в США, где Amphorophora agathonica (?) иногда изредка встречается.
Ниже перечислены в порядке уменьшения их значимости в качестве переносчиков в Северной Америке следующие виды тлей, переносящие компонентов комплекса мозаики малины: Amphorophora agathonica (?) Hottes [11], Masonaphis (Oestlundia) rubicola (Oestlund [13], Amphorophora sensoriata Mason [13] и A. rubi-cumberlandi Knowlton et Allen [138].
Было установлено, что Amphorophora rubitoxica Knowlton [217], A. rubi (Kalt.) subsp. ruborum Born., которые встречаются на ежевике в Европе [36], Aphis rubicola Oestlund [11], A. idaei van der Goot [40] и Chaetosiphon fragaefolii (Cockerell) [229] не являются переносчиками
Amphorophora rubi (Kalt.) subsp. idaei Born, (обычно цитируемая как A. rubi) обнаружена на дикой и культурной малине (Rubus idaeus ;L.) в Европе, но была отмечена на сорте Ллойд Джордж в Саммерленде, Британская Колумбия (Стейс-Смит; не опубликовано).
Биология A. rubi изучалась Дикером [88].
Amphorophora agathonica (?) широко распространена в Северной Америке на дикой и культурной малине и на малине японской dRubus phoenicolasius Maxim.) на востоке США [63]. Бойзенберри [72], некоторые сорта ежевики [261, Конверс и Раселл; не опубликовано] и, возможно, виды Fragaria [159] являются зарегистрированными хозяевами, но тля в общем ограничивается красной и японской малиной. Биология A. agathonica (?) исследовалась Винтером [261]. Обзор о таксономии тлей, встречающихся на DRubus в Северной Америке, был сделан Палмером [186]. Идентификация североамериканской популяции "A. rubi" как A. agathonica (?) была предложена Кеннеди и др. [155]. (См. также примечание в начале этой статьи.) Однако почти вся литература до 1967 г. относит обычную североамериканскую большую малинную тлю к Amphorophora rubi.
Среди сортов малины имеются значительные различия в их способности поддерживать колонии тлей (см. стр. 186). В части статей об устойчивости красной малины к заселению британской популяцией A. rubi Бриггс [28, 29, 30] описал методы оценки этой устойчивости к заселению в тепличных и полевых условиях. Он перечислил четыре линии A. rubi в Англии, различающиеся по своей способности к заселению определенных дифференцирующих сортов малины с известным генотипом. Бриггс полагает, что генетический контроль реакции хозяина на эти четыре линии осуществляется двумя парами аллелей. Несмотря на их способность заселять хозяев, несущих антиколонизационные гены A1, А2, А3, линия № 4 A. rubi отличается низкой врожденной плодовитостью и поэтому не составляет угрозы для восприимчивых сортов в Англии. Старые желтые листья устойчивых сортов иногда заселялись A. rubi [28, 160].
Amphorophora Agathonica (?) (Северная Америка) и A. rubi (Европа) заметно различаются по способности заселять определенные сорта малины [88, 127] (см. стр. 186).
Распространение в естественных условиях. В условиях промышленного размножения посадочного материала зараженные саженцы являются основной причиной распространения вирусов [64], поэтому основные способы борьбы должны быть сосредоточены на выращивании здорового посадочного материала. Мозаика может также распространяться вдоль ряда от инфицированного растения, когда приживутся его отпрыски или окоренившиеся верхушки.
Изучалась скорость передвижения вирусов мозаики внутри инокулированных растений малины. Беннетт [13] установил в опытах с кольцеванием, что ТЛКМ передвигается вниз в побегах черной малины примерно со скоростью 5 мм в час; у ТС КМ скорость движения медленнее, но ни один из вирусов этих групп не передвигался через окольцованный участок стебля. Фримен и Стейс-Смит [97] прививали культуры ТЛКМ и ТСКМ в побеги красной малины в августе и нашли, что культура ТЛКМ обнаруживалась в корневых черенках, взятых 4 месяца спустя, с частотой, почти в 2 раза большей, чем культура ТСКМ. Они нашли, что ни один из вирусов не передвигался в течение всего периода, когда растения находились в состоянии покоя. Беннетт [13] подчеркивал, что разная скорость передвижения этих двух групп вирусов в растениях малины благоприятствует экспериментальному разделению их из смеси, встречающейся в естественных условиях.
Распространение мозаики с помощью переносчика, ограниченное Amphorophora rubi для Европы и A. agathonica (?) для Северной Америки, происходит двумя способами: относительно большой дальностью перелета крылатых тлей и местным распространением бескрылыми тлями. Крылатые особи встречаются в Англии в июне или начале июля [88]. Кадмен (не опубликовано) установил, что период миграции малинных тлей длится с конца июня до середины августа. Их максимальный инфекционный радиус действия не известен, но в США и Англии имеются полевые данные о том, что инфекционный радиус крылатых насекомых A. agathonica (?) и A. rubi соответственно должен быть больше 1/4 мили и, вероятно, гораздо более (Кадмен, не опубликовано; 67; Стейс-Смит, не опубликовано).
Местное распространение. Местное распространение мозаики в поле, вероятно, осуществляется в первую очередь бескрылыми тлями. Хотя бескрылые особи могут медленно передвигаться от растения к растению вдоль ряда или между рядами, представляется вероятным, что ветер, дождь и междурядная обработка являются важными факторами их перемещения [261].
Пик популяций тлей Amphorophora rubi развивается в Англии в середине лета и еще раз в начале осени [88], а в Нидерландах обычно в конце июля [160]. Максимум инвазии тлями в Британской Колумбии обычно наблюдается в конце июня [224]. В штатах Мичиган и Вашингтон самые крупные популяции Л. agathonica (?) были обнаружены в конце июля и начале августа [13, 141], и они увеличивались в течение всего лета в Вермонте [Конверс и др., не опубликовано]. Мозаика распространяется радиально от отдельного источника [13], но часто продвигается быстрее вдоль рядов, чем поперек их [76, 196]. Скорость распространения значительно варьирует в различных районах.
Ранкин и его коллеги [76, 194, 195, 196] проводили обширные исследования по скорости распространения мозаики в поле, но основывались только на полевых симптомах и поэтому спутали новые инфекции с более ранними латентными инфекциями, которые стали очевидными. В полевых условиях уровень видимой мозаики был между 0 и 2% в начале наблюдений, а спустя 12 месяцев он варьировал от 10 до 28%. На плантации черной малины в штате Нью-Йорк, где не удалялись больные растения, видимая мозаика достигала в 1927 г. 27%, в 1928 г. - 47% и в 1929 г. - 53% [196]. Сообщалось, что ежегодная скорость распространения видимой мозаики вдоль ряда составляла от 1,5 до 13,8 м [194]. Кадмен и Фис-кен [48] и Кадмен [40] изучали скорость распространения ТЛКМ и ТСКМ в Шотландии, где ТЛКМ распространялся в поле быстрее, чем ТСКМ. Согласно сообщениям, без проведения борьбы с тлями скорость заражения ТЛКМ в естественных условиях в течение одного года была равна 60%.
В США Конверс [67] изучал скорость распространения мозаики (главным образом ТЛКМ) в свободном от нее посадочном материале на 14 полевых участках в 9 восточных ,и среднезападных штатах в течение двух лет. Скорость распространения варьировала от 0% в девяти местностях до максимум 7% на одном участке в штате Нью-Джерси. Скорость распространения инфекции мозаики на девяти сортах малины в среднем по всем участкам колебалась от 0 до 10%. Степень изоляции от диких, и культивируемых Rubus и интенсивность применения инсектицидов в борьбе с тлями, по-видимому, были не так важны, как сорт в определении процента заражения мозаикой.
Меры борьбы. Для уменьшения распространения заболевания в посадочном материале существует четыре способа: 1) уничтожение зараженных культивируемых и диких видов растений - источников заболевания; 2) химическая борьба с тлями-переносчиками; 3) селекция на устойчивость хозяина к заселению тлями; 4) селекция на устойчивость хозяина к вирусам мозаики.
1. Уничтожение зараженных культивируемых и диких растений-источников заболевания. В большинстве районов выращивания малины человек в значительной мере ответствен за распространение мозаики. Многие из основных сортов Rubus, особенно красной малины на промышленных плантациях, обычно заражены ТЛКМ или ТЛКМ и ТСКМ [37, 64, 220]. Одно время удаление больных растений с участка являлось главным способом борьбы с мозаикой в питомниках малины. Хотя тщательное удаление ценно для уменьшения видимой мозаики [149, 194, 203], латентные вирусы мозаики остаются необнаруженными и снижают энергию роста и урожайность. Такие инфицированные растения подвержены более серьезному поражению, когда их заражают добавочные вирусы мозаики, и множественная инфекция дает вспышку мозаики малины.
Общая дискуссия о роли инфексации и сертификации в борьбе с мозаикой приведена во введении (см. стр. 169).
Практикуется еще удаление больных растений на плодоносящей плантации малины, но только в качестве средства продления продуктивной жизни плантации. Однако основным способом борьбы с мозаикой малины является посадка свободных от мозаики клонов на некотором расстоянии от зараженных растений Rubus.
Дикорастущая красная малина - важный резерватор мозаики. Переносчики тли всегда присутствуют, где бы она ни находилась [40, 63, 77]. На востоке штатов США японская малина (Rubus phoenicolasius Maxim.) также обычно заражена мозаикой и служит убежищем для тлей-переносчиков мозаики [63]. Уничтожение диких растений малины до посадки культивируемых видов Rubus снижает появление видимой мозаики на выращиваемых сортах [77]. Для уничтожения растений Rubus имеются эффективные гербициды [1], но для предотвращения восстановления дикой малины из семян требуются повторные обработки.
Минимальная изоляция, требуемая для отделения культивируемых видов Rubus от зараженных мозаикой диких растений сложно-плодных, не может быть точно определена. Рекомендуются расстояния примерно 400, 200 и 100 м [5, 23, 40, 67, 260].
2. Химическая борьба с тлями-переносчиками. Имеются несколько инсектицидов, которые эффективны против Amphorophora rubi и A. agathonica (?) в течение нескольких недель после опрыскивания. Некоторые из них являются для растения системными [48; Шеферс, не опубликовано]. Кадмен (не опубликовано) установил, что даже при частом применении системных инсектицидов в однолетком опыте в Шотландии 20% растений красной малины были заражены мозаикой по сравнению с 60% на неопрыснутых участках. Передача вирусов полуперсистентного типа, например вируса мозаики малины, может быть выполнена одним проколом ткани стилетом одной тли, даже если тля погибнет сразу после этого. Поэтому заражение плантаций вирусами, переносимыми тлями, не может быть полностью предотвращено с помощью инсектицидов [58]. Однако предотвращение заселения вирофорными Amphorophora с помощью инсектицидов уменьшает или предотвращает повторное распространение инфекции. Следовательно, изоляция плантаций ягодных культур от источников вирофорных тлей повышает эффективность инсектицидов в борьбе с мозаикой.
3. Селекция на устойчивость хозяина к заселению тлями. Ранкин [194] впервые отметил, что среди сортов малины наблюдаются заметные различия в восприимчивости к мозаике, и предложил термин "klendusity" для обозначения устойчивости к заражению или способности избегать его. Однако этот термин является слишком широким и неопределенным, чтобы быть полезным. Наследственная устойчивость к питанию и заселению тлей Amphorophora agathonica (?) впервые была доказана в США [141, 206]. Хубер и Шварц указали на присутствие двух доминантных генов. Добени [84] показал, что иммунитет к тле A. agathonica (?) в Британской Колумбии у сортов красной малины, происшедших от сорта Ллойд Джордж, обусловлен одним доминантным геном. Согласно Конверсу и Бейли [72], Добени и Стейс-Смиту [85], английский сорт красной малины Ллойд Джордж уже успешно использовался в селекции для придания стабильной устойчивости к тле A. agathonica (?) ряду американских сортов. Однако не все американские сорта, имеющие в родословной сорт Ллойд Джордж, обладают устойчивостью [72].
Классическая работа Найта и др. [157, 158] указывает на присутствие семи аллельных генов у английского сорта Сеянец "А" Бомфорта и американского сорта красной малины Чиф, придающих устойчивость к четырем английским расам тли Amphorophora rubi. Найт и др. [158] нашли, что ген устойчивости А1 к тле был сцеплен с нормальным аллелем полулетального гена. В ФРГ Баумайстер [9, 10] также изучал генетику устойчивости к A. rubi. Селекционные программы по введению генов устойчивости к переносчикам проводятся на Ист-Моллингской станции [157], в Данди [146] и в Швеции [185]. В Канаде [84] и в США возобновился интерес к такого рода работе.
Несколько сортов красной малины необычны в своем взаимодействии с Amphorophora agathonica (?) и мозаикой малины. Стейс-Смит [224] показал, что растения сорта красной малины Ренер, инфицированные мозаикой, могут поддерживать большие колонии A. agathonica (?), но последние не в состоянии получать из них вирус. В противоположность этому, сорт Септембер поддерживает большие колонии A. agathonica (?), но редко поражается мозаикой в полевых условиях и не заражается экспериментально при последующем заселении крупными популяциями вирофорных тлей. Однако он легко заражался прививкой, после чего тли легко получали вирус мозаики (ТЛКМ) и переносили его на восприимчивых хозяев (Конверс, не опубликовано).
4. Селекция на устойчивость растений-хозяев к вирусам мозаики. В конечном счете есть возможность вести борьбу путем использования устойчивости хозяина к заражению вирусами мозаики малины или против проявления болезней, которые они вызывают [45, 146]. По-видимому, английский сорт Норфолк Джайент избегает заражения мозаикой в Англии, хотя он заселяется переносчиком [48]. Так же ведет себя сорт Уилламетт на северо-западном побережье Тихого океана в Северной Америке [220]. Это примеры скорее низкой восприимчивости, а не абсолютного иммунитета к заражению мозаикой. Однако еще не ясно, представляют ли приведенные сорта в примерах устойчивость к очень ранним стадиям вирусной инфекции после передачи переносчиком или устойчивость в отсутствии насекомого-переносчика.
Учитывая ограниченность источников генов устойчивости малины к Amphorophora rubi или к вирусам мозаики, английские селекционеры пытаются идентифицировать и включать в промышленные сорта оба вида устойчивости.
Терапия. Стейс-Смит [223] установил, что вирусы мозаики малины делятся на две группы: вирусы, сохраняющиеся в хозяине в течение 3 месяцев или более при постоянной температуре 37°С, и вирусы, инактивируемые в растениях-хозяевах при выращивании от 5 до 10 дней при 37°С. Вирус, представляющий ТЛКМ, сохранял половину инфекционности после обработки малины в течение 62 ч при38±1,5°С [70].
В последующем работа Чамберса [61] по термотерапии была успешно использована для получения свободного от вирусов посадочного материала многих сортов малины [20, 61, 67, 69, 70, 177] и некоторых сортов ежевики (Конверс, не опубликовано).
Примечания. Исключительное разнообразие симптомов на малине при поражении вирусами мозаики отмечали многие исследователи [И, 13, 45, 70, 76, 108, 110, 195, 218]. Мы можем предполагать, что внутри двух групп ТЛКМ и ТСКМ имеется большое число штаммов или отдельных вирусов. Изменчивость в выражении симптомов, возникающих от одного полевого источника мозаики малины, является загадкой. Кули [76] показал, что передача инфекции с растений пурпуровой малины сорта Колумбийская, отличающихся однородностью в образовании симптомов, на черной малине вызывала симптомы мозаики широко различающихся типов. Конверс [70] установил, что непрогретые растения сорта Латам с симптомами и без них одинаково вызывали сильные симптомы, когда использовались в качестве инокулюма, на Rubus henryi и R. occidentalis. В настоящее время нет соответствующих методов исследования вирулентности ТЛКМ или ТСКМ или определения их константности.

Рис. 65. Некротическая реакция верхушки побега Rubus henryi, инокулированной прививкой листочком одним из термолабильных компонентов вируса мозаики
Обнаружение и идентификация. Любой из вирусов, который при передаче его на R. henryi прививкой будет вызывать некроз сильно растущих верхушек или боковых почек, может быть отнесен к компонентам вирусов мозаики малины. Если этот вирус может быть передан между растениями-хозяевами Rubus с помощью тлей Amphorophora rubi или A. agathonica (?) и в последующем будет вызывать некроз у R. henryi (рис. 65), крапчатость, посветление жилок или некроз на R. occidentalis, его относят к группе вирусов мозаики малины. Тепловая обработка хозяина в течение одной недели при 37°С позволит отнести его к группе ТЛКМ или ТСКМ. В Европе сорта Сайнт Вальфрид, Норфолк Джайент, Сеянец "Б" Бомфорта и Моллинг Ландмарк используются для идентификации обнаруженных специфичных мозаичных вирусных заболеваний [35; Кадмен, не опубликовано].
ЖИЛКОВЫЙ ХЛОРОЗ МАЛИНЫ (RASPBERRY VEIN CHLOROSIS)
Синонимы. Raspberry chlorotic net [150]. История и географическое распространение. Болезнь впервые была описана в Шотландии Кадменом [32]. Однако данные о передаче тлей Aphis idaei van der Goot были опубликованы раньше [51]. Эти ранние сообщения были подтверждены и расширены Стейс-Смитом [225]. Это заболевание является обычным на культивируемых и дикорастущих растениях малины (Rubus idaeus L.) в Англии. Наблюдения и сообщения, так же как и исследования Иордовича [150], наводят на мысль, что болезнь широко распространена в Западной Европе.
Она встречается в Канаде [225] и Новой Зеландии (Кадмен, не опубликовано), но не было сообщений из других стран, выращивающих малину, таких, как США и Австралия.
Экономическое значение. Не известно.
Круг растений-хозяев. По-видимому, естественным хозяином является красная малина, и обычно все выращиваемые европейские сорта восприимчивы к заражению. Некоторые североамериканские сорта (Ньюбург, Кутберт, Латам и Викинг), по-видимому, иммунны [32], тогда как другие, например Вашингтон, восприимчивы [225]. Ни вирус, ни симптомы не обнаружены на культивируемых или диких видах Rubus, кроме R. idaeus. Однако вирус был перенесен прививкой на логанову ягоду (R. loganobaccus Bailey) и с помощью тлей (Aphis idaei) на альпийскую землянику (Fraga-ria vesca var. semperflorens) [225]. В опытах с прививками Rubus henryi Hemsl. et. Kuntze и R. occidentalis L. не заразились [225].
Симптомы. На красной малине самые мелкие жилки на листьях становятся желтыми или желтовато-зелеными, и на листьях образуется желтый сетчатый узор, похожий на узор, вызываемый вирусом желтой сетчатости малины, но более резко выраженный. На некоторых сортах, например Ллойд Джордж, хлороз локализованный и пятнистый, а листья могут быть деформированными (рис. 66).
![Рис. 66. Слева - лист сорта красной малины Ллойд Джордж через 5 недель после инокуляции тлей; справа - на том же сорте проявляется обширный хлороз, который виден на некоторых листьях зараженного растения [225]](pic/000072.jpg)
Рис. 66. Слева - лист сорта красной малины Ллойд Джордж через 5 недель после инокуляции тлей; справа - на том же сорте проявляется обширный хлороз, который виден на некоторых листьях зараженного растения [225]
Три типа симптомов - слабые, средние и сильные - различимы на красной малине, и, очевидно, они вызываются родственными штаммами вируса [32].
Вирус хлороза жилок вызывает посветление и искривление жилок при экспериментальном заражении логановой ягоды и альпийской земляники.
Характеристика вируса. Вирус не передавался инокуляцией сока на травянистые растения. Нет сведений о его свойствах.
Передача. Вирус передается прививкой и тлями Aphis idaei, но не Amphorophora rubi (Kalt.) [32, 51] или A. agathonica (?) Hot-tes [225]. Экспериментально A. idaei является неэффективным переносчиком, и трудно получить соответствующие результаты. Имеются указания, что хлороз жилок вызывается вирусом циркулятивного типа; тлям для накопления вируса требуется период питания по крайней мере 24 ч и вирус сохраняется в них в течение 24 ч. В этом отношении взаимосвязь между переносчиком и ВЖХМ отличается от взаимодействия вирусов, переносимых тлями A. rubi и A. agathonica (?).
Распространение в естественных условиях. Нет оснований сомневаться в том, что Aphis idaei является естественным переносчиком. В Англии популяции тли по численности резко меняются год от года. Согласно полевым наблюдениям, вирус распространяется медленно, но нет точных данных.
Меры борьбы. В практике удаление больных растений обеспечивает борьбу с распространением заболевания в питомнике малины.
Терапия. По-видимому, вирус термостабилен in vitro, так как больные растения не освобождались от инфекции, когда содержались при 37°С в течение 3 месяцев [223] или при 40°С в течение 60 дней [62].
Обнаружение и идентификация. Характерные симптомы на стандартных индикаторных сортах, например Норфолк Джайент или Ллойд Джордж, зараженных прививкой или тлями, являются единственным критерием для идентификации.
КУРЧАВОСТЬ ЛИСТЬЕВ МАЛИНЫ (RASPBERRY LEAF CURL)
Синонимы. (Сокращенно RLCV.) Curl, raspberry curl, yellows. Последний синоним был ранним обозначением, которое обычно включало мозаику. Шотландская курчавость листьев малины - совершенно другое заболевание, вызываемое вирусом кольцевой пятнистости малины [38].
История и географическое распространение. Курчавость листьев была одной из первых определенных вирусных болезней сложно-плодных. Рое [202] отмечал, что курчавость листьев является серьезной болезнью красной малины в штате Нью-Йорк, которая могла распространяться от растения к растению. Ранкин и Хокей [197] предполагали, что вирус передавался Aphis rubiphila Patch (= A. rubicola Oestlund). Смит [213] и Беннетт [11] подтвердили это подозрение.
Беннетт установил существование двух типов вируса курчавости, которые он назвал альфа и бетта вирусы курчавости. Оба вируса известны только в Северной Америке. Вирус альфа поражает красную и пурпуровую малину, а вирус типа бета заражает также и черную малину. Оба типа вируса могут заражать ежевику, но это наблюдается редко. Курчавость листьев малины встречается в США и Канаде всюду, где возделывается малина, за исключением зоны северо-западного побережья Тихого океана, что объясняется отсутствием переносчика тли Л. rubicola в этой зоне. Курчавость редко встречается к югу от Нью-Йорка в восточных штатах США, но имеет важное значение на красной малине в Новой Англии и районе Скалистых гор и на черной малине в штатах Мичиган и Огайо.
Экономическое значение. Курчавость листьев малины можно считать вторым заболеванием после мозаики, составляющим проблему вирусов малины в Северной Америке. Урожай красной малины снижается на 20 - 40% по сравнению со здоровыми растениями; зараженные растения, особенно черной малины, могут отмирать после нескольких сезонов [11].
Круг растений-хозяев.
Естественные хозяева. Rubus, подрод Idaeobatus. R. idaeus L. var. strigosus (Michx.) Maxim (красная малина), вирус альфа или бета. Сорт Кутберт - хорошее тест-растение R. occidentalis L. (черная малина), только вирус бета R. neglectus Peck (гибриды пурпуровой малины), вирус альфа или бета R. phoenicolasius Maxim, (малина японская), вирус альфа или бета. Rubus, подрод Eubatus (растения-хозяева меньшего значения) R. allegheniensis Porter (лесная ежевика) и сорт Эльдорадо, вирус альфа и бета R. procerus P. J. Muell. (ежевика Гималайя) R. ursinus Cham, et Schlecht. (стелющаяся ежевика Тихоокеанского побережья) Экспериментальные хозяева R. albescens Roxb. (тропическая черная малина), вирус бета (Конверс; не опубликовано) R. baileyanus BrittonXfl. argutus Link (сорт Лукреция) [11] R. henryi Hemsl. et Kuntze, вирус альфа и бета [226, Конверс; не опубликовано] Fragaria vesca L., вирус альфа [226]
![Рис. 67. Вверху - начальные симптомы поражения вирусом альфа курчавости листьев на красной малине сорта Ллойд Джордж; внизу - хронические симптомы на том же сорте [226]](pic/000073.jpg)
Рис. 67. Вверху - начальные симптомы поражения вирусом альфа курчавости листьев на красной малине сорта Ллойд Джордж; внизу - хронические симптомы на том же сорте [226]
![Рис. 68. Вверху - хронические симптомы поражения вирусом бета курчавости листьев на черной малине сорта Нью Логан. Внизу - симптомы вируса альфа курчавости листьев на сеянце японской малины (Rubus phoenicolasius) [226]](pic/000074.jpg)
Рис. 68. Вверху - хронические симптомы поражения вирусом бета курчавости листьев на черной малине сорта Нью Логан. Внизу - симптомы вируса альфа курчавости листьев на сеянце японской малины (Rubus phoenicolasius) [226]

Рис. 69. Симптомы курчавости листьев на Rubus henryi
Симптомы.На культурных растениях-хозяевах.Красная малина. Растения без симптомов или проявляют большей частью слабое закручивание вниз верхушечных листьев в год заражения (рис. 67, вверху). Следующей весной листья на плодоносящих и однолетних побегах курчавые, слегка желтоватые (рис. 67, внизу). Плодоносящие боковые побеги укорочены, и может наблюдаться пролиферация побегов в виде розетки. Новые побеги карликовые, многочисленные и ветвящиеся из пазух листьев. Растения остаются карликовыми и часто погибают в течение следующей зимы. Ягоды на больных растениях мелкие и рассыпаются.
Черная малина. Симптомы такие же, как на красной малине. Листья дугообразно изогнуты, плотные, остаются мелкими и почти округлыми по очертанию, темно-зелеными с маслянистым оттенком (рис. 68, вверху). При хронической инфекции молодые побеги негибкие, хрупкие и часто не ветвятся.
Ежевика. На некоторых сортах ежевики видны симптомы курчавости листьев, как на красной малине, тогда как другие сорта остаются бессимптомными.
На индикаторных растениях.Японская малина. Симптомы обычно видны через 7 - 10 дней после инокуляции тлями. Признаки заболевания такие же, как на красной малине, но значительно более ярко выражены. Черешки верхушечных листьев закручены вниз. Листовая пластинка не развертывается нормально, и межжилковая ткань в неразвернутой части хлоротичная. Последующие листья курчавые и мелкие, образуют розетку на верхушке растения (рис. 68, внизу).
Rubus henryi. Первые симптомы заметны через 10 - 14 дней после инокуляции тлями. Молодые листья хлоротичны и асимметрично изогнуты (рис. 69). В течение месяца после инокуляции верхушки побегов и пазушные почки становятся некротическими.
Альпийская земляника. Межжилковый хлороз заметен через три недели после инокуляции прививкой. Этот хлороз виден на двух или трех последующих листьях; позднее инфицированные растения склонны к выздоровлению и не отличаются от контрольных растений.
Присутствие латентного штамма А вируса морщинистости земляники на индикаторном клоне не усиливает симптомы, как это происходит с некоторыми вирусами земляники (см. стр. 35).
Характеристика вируса. Штамм альфа вируса курчавости малины не проявляет перекрестной защиты против штамма бета [12], поэтому мнение, что альфа и бета являются штаммами одного вируса, сомнительно. Альфа вирус курчавости часто не обнаруживается в зараженных растениях через несколько месяцев после инокуляции. Такие растения могут быть повторно заражены и, таким образом, будут, очевидно, спонтанно выздоровевшими, что представляет редкое явление в растительной вирусологии [12]. Передвижение альфа вируса курчавости в растении малины ограничено флоэмой и относительно медленное [И]. Другие сведения о морфологии, свойствах вируса и серологических взаимоотношениях отсутствуют.
Передача. Вирус курчавости листьев малины передается прививкой пластинкой коры и черешком в расщеп так же хорошо, как и тлями [109, 213, 226]. Обычным переносчиком является Aphis rubicola Oestlund. Малая малинная тля A. rubifolii (Thos.) может инфицировать ежевику, но положительные данные получены только в полевых опытах [11]. Aphis idaei van der Goot - экспериментальный переносчик альфа вируса курчавости [226]. Сортовая устойчивость к тлям-переносчикам не известна. Данные о переносчиках приведены в таблице 8. Вирус, отмеченный Боннмезоном [21] во Франции, который он переносил с помощью тлей A. idaei, был, вероятно, возбудителем жилкового хлороза.
Было найдено, что Amphorophora agathonica (?) Hottes (указываемая в литературе как A. rubi), A. sensoriata Mason, Macrosiphum (Amphorophora) rubicola (Oestlund) и Aphis spiraecola не являются переносчиками [12, 65, 226].
Распространение в естественных условиях. В естественных условиях вирус распространяется тлей Aphis rubicola Oestlund. Бескрылые тли медлительны и имеют небольшое значение в распространении вируса. Крылатые тли, которые развиваются в июне, могут переноситься токами воздуха на значительные расстояния.
Меры борьбы. Поскольку бескрылые особи A. rubicola сравнительно малоподвижны, осмотры и уничтожение больных кустов являются эффективными мерами борьбы. Там, где заболевание очень сильное, желательно уничтожение находящихся вблизи растений дикой малины, которые служат убежищем для вируса и переносчика. В таких случаях применение инсектицидов для уменьшения популяций переносчиков на плантациях также может быть полезным. Против штаммов альфа и бета вируса курчавости существует иммунитет [65], но, к сожалению, не было сделано попыток вывести генетически иммунные или устойчивые сорта.
Терапия. Альфа вирус курчавости не был уничтожен в зараженных растениях, содержавшихся при 37°С до четырех недель [226]. Однако способ термотерапии еще разрабатывается, поскольку нет сортов, которые были бы полностью зараженными. Как указывалось выше, были отмечены случаи спонтанного выздоровления растений при полевых температурах.
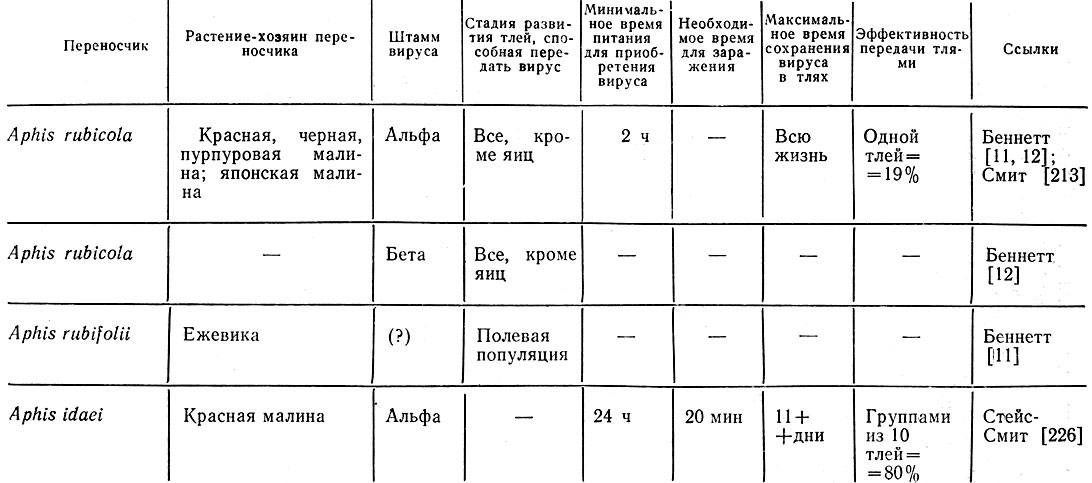
Таблица 8. Взаимоотношения переносчик - вирус - растение-хозяин при курчавости листьев малины
Обнаружение и идентификация. Эти вирусные болезни могут быть легко обнаружены в любое время вегетационного периода по плотной курчавой листве на больных растениях. Однако нужно иметь в виду, что заселение тлями Aphis rubifolii может вызывать сильное закручивание листвы растений ежевики при отсутствии курчавости [137]. Очень обильные популяции A. rubicola, часто находящиеся на листве молодых растений черной малины, также могут вызывать обманчивые симптомы. Хронически инфицированные растения малины всегда сильно отстают в росте, слабо ветвятся и имеют курчавую листву. Малина японская - хорошее индикаторное растение-хозяин для латентной инфекции, но как черная, так и красная малина должны использоваться, чтобы отличить между собой альфа и бета штаммы вируса курчавости.
ВИРУС ОГУРЕЧНОЙ МОЗАИКИ НА МАЛИНЕ
Синонимы. Нет. (Сокращенно CMV.)
История и географическое распространение. Заболевание описано в 1958 г. и известно только на нескольких плантациях красной малины (Rubus idaeus L., сорт Ллойд Джордж) в восточной Шотландии [117]. Однако Цундель [270] сообщил, что растения черной малины, росшие вблизи плантаций тыкв в Пенсильвании, проявляли симптом папоротниковидное™ листа, и предположил, что это могло быть вызвано вирусом огуречной мозаики.
Экономическое значение. Небольшое или не имеет. При осмотре плантаций случаи поражения составили, вероятно, менее 1%.
Круг растений-хозяев. Вирус огуречной мозаики имеет широкий круг хозяев как среди сельскохозяйственных и декоративных растений, так и среди травянистых видов, выращиваемых в теплице в качестве тест-растений для определения вирусов. Среди видов ягодных культур сообщалось также об этом вирусе на черной [243] и красной смородине. Черная малина (R. occidentalis L.) может быть заражена экспериментально. Нет сведений о реакции других сортов малины, кроме Ллойд Джордж, или других видов Rubus, кроме Rubus idaeus или R. occidentalis, на инокуляцию инфекционных препаратов прививкой или с помощью переносчиков.
Симптомы. а) На малине. Симптомы известны только на растениях сорта Ллойд Джордж, зараженных также неописанным латентным вирусом (Кадмен, не опубликовано). Не известно, влиял ли латентный вирус на симптомы, вызываемые вирусом огуречной мозаики.
Симптомы состояли из светло-зеленых крапинок, окруженных темно-зелеными участками нормального цвета (рис. 70). При беглом осмотре не отмечено влияния заболевания на рост, образование отпрысков или урожай ягод.

Рис. 70. Лист красной малины сорта Ллойд Джордж, пораженный в естественных условиях вирусом огуречной мозаики.
б) На индикаторных растениях-хозяевах. Ниже перечислены несколько пригодных индикаторов.
Табак (Nicotiana tabacum L., сорт Уайт Барлей).На инокулированных листьях обнаруживаются хлоротичные поражения или симптомы отсутствуют; на системно зараженных листьях развивается зеленая мозаика.
Огурец (Cucumis sativus L., сорт Эвридей). На инокулированных листьях обнаруживаются хлоротичные поражения; на системно зараженных листьях развивается светло- и темно-зеленая мозаика.
Карликовая фасоль (Phaseolus vulgar is L., сорт Принц). Зимой на инокулированных листьях обнаруживаются поражения в виде бурых точек в течение двух дней при 20° С, но в Англии летом симптомов не бывает. Вирус редко может быть обнаружен в инокулированных листьях. Болезнь не является системной на этом растении.
Chenopodium amaranticolor Coste et Reyn.; С. quinoa Willd. На инокулированных листьях видны хлоротичные поражения с небольшими некротическими центрами. Инфекция несистемная.
Nicotiana clevelandii Gray, N. glutinosa L., Datura stramonium L. и Petunia hybrida Vilm. проявляют системные мозаичные симптомы.
Растения томата (Lycopersicum esculentum Mill.) становятся системно пораженными и образуют узкие листочки - симптом папоротниковидности листьев.
в) Симптомы при сочетании с другими вирусами. За исключением описанных выше симптомов на малине, о каких-либо других не сообщалось.
Характеристика вируса. Типичные штаммы вируса огуречной мозаики, подобно штаммам, преобладающим на других естественно зараженных растениях в Англии, вызывают симптомы зеленой мозаики на растениях табака. Свойства вируса в соке табака следующие: температура инактивации колеблется между 62 и 68° С в течение 10 мин; предельное разведение находится между 10-3 и 10-4; сохранение инфекционности при 20°С - от 3 до 5 дней.
Изоляты вируса из растений малины не были очищены и изучены серологически. Изоляты с других растений-хозяев могут быть очищены, согласно описанию Скота [207]. Они содержат примерно 18% рибонуклеиновой кислоты [153] и имеют изометрические частицы диаметром около 30 ммк, которые в электронном микроскопе на напыленных препаратах представляются уплощенными. При обработке фосфоровольфрамовой кислотой частицы относительно мало контрастны и без предварительной фиксации (2%-ным раствором формальдегида) склонны к дезинтеграции. Их коэффициент осаждения при неограниченном разведении водой при 20° С равен примерно 100 S.
Вирус осаждается 0,85%-ным хлористым натрием [94], который поэтому не должен использоваться в качестве разбавителя при серологических преципитиновых тестах, проводимых в смешанных жидкостях. Его серологическое поведение описано детально Скоттом [208].
Передача. а) Способы. 1. Прививкой. По-видимому, возможна, но никогда не испытывалась на малине.
2. Повиликой. Никогда не испытывалась на малине, но возможна на других хозяевах с помощью ряда видов Cuscuta [205].
3. Семенами и пыльцой. Не испытывалась на малине. Сообщалось о передаче семенами у других видов.
4. Инокуляцией сока. Вирус с трудом передавался с малины на травянистые растения. Передача улучшается при растирании зараженных листьев в среде, содержащей 0,01М диэтилдитиокарбамат и 0,1М фосфорный буфер (рН 8,5) или 2,5% никотина, для снижения действия инактиваторов вируса и ингибиторов инфекции, имеющихся в соке из листьев малины. Нет сведений о передаче с травянистых растений на малину и с малины на малину. Вирус легко передается с одного травянистого растения (например, табака) на другое.
5. Переносчики. Многие тли являются переносчиками вируса [155]. Amphorophora rubi (Kalt.) и Myzus persicae (Sulz.) передавали изоляты вируса, первоначально полученные с растений малины; A. rubi, но не М. persicae, передавали вирус с Datura stramonium на Rubus occidentalism никакие другие виды не передавали его с малины на D. stramonium [117].
б) Взаимоотношения вирус - переносчик. Изоляты вируса из растений малины не испытывались. Другие изоляты передавались тлями по неперсистентному типу, как показано здесь.
Пороговые периоды приобретения вируса и инокуляции: менее 1 мин.
Сохранение способности передавать инфекцию: при питании менее 1 ч, при голодании менее 8 ч. Латентного периода в переносчике нет.
На молодых растениях табака через неделю после инокуляции тлями обнаруживаются симптомы системного заражения.
Распространение в естественных условиях. По-видимому, вирус распространяется по плантации малины тлями-переносчиками. Однако зараженные растения встречаются в разных местах плантации, и это говорит о том, что мало или нет повторного распространения вируса тлями-переносчиками от растения к растению. Зараженные побеги как посадочный материал могут быть разносчиками заболевания. Нет данных о распространении вируса посредством приемов агротехники.
Меры борьбы. Способы борьбы не проверялись, но удаление больных растений должно быть эффективным.
Терапия. На малине не испытывалась. Термотерапия против некоторых штаммов вируса является успешной на других видах растений.
Примечания. По-видимому, заражение малины вирусом огуречной мозаики представляется необычным и несущественным. Нет данных о том, что малина служит резерватором вируса для других видов растений.
Обнаружение и идентификация. а) Обнаружение. Заражение осуществляется инокуляцией сока малины на Chenopodium quinоа или С. amaranticolor.
б) Идентификация. Она проводится путем серологических тестов в агаровом геле методом двойной диффузии. Проводятся также тесты перекрестной защиты, для того чтобы определить способность неидентифицированного изолята вируса защитить системно зараженные растения табака от инокуляции тест-изолятом вируса огуречной мозаики, вызывающим некрозы на инокулированных листьях растений табака, свободных от вируса.
ВИРУСНАЯ КОЛЬЦЕВАЯ ПЯТНИСТОСТЬ МАЛИНЫ ДУШИСТОЙ (THIMBLEBERRY RINGSPOT)
Синонимы. Нет.
История и географическое распространение. Эта болезнь была описана Стейс-Смитом [222]. Она встречается в естественных условиях на дикой малине душистой (Rubus parviflorus Nutt.) и ограничена районами нижней Фрезер Валли в Британской Колумбии.
Экономического значения не имеет.
Круг растений-хозяев. Естественным хозяином является душистая малина. Экспериментально вирус был передан на черную малину (R. occindentalis L.), красную малину (R. idaeus L.) и R. hen-ryi liemsl. et Kuntze.
Симптомы.Красная малина (сорт Вашингтон). На листьях виден слабый хлоротичный рисунок дубового листа или кольцевая пятнистость (рис. 71, вверху слева). Симптомы более выражены весной, чем летом.
Черная малина (сорт Мунгер). На листьях проявляется хлоротичная крапчатость, иногда кольцевая пятнистость (рис. 71, вверху справа). Верхушки стебля погибают, и пробуждаются пазушные почки, придавая растению кустистый, розеточный тип роста.
Душистая малина. На листьях виден неправильный рисунок кольцевой пятнистости. Кольца мелкие и неотчетливые на молодых листьях, но более крупные и четко выраженные на старых листьях (рис. 71, внизу слева). Наблюдается значительная изменчивость интенсивности симптомов от растения к растению.
Rubus hеnrуi. На молодых распустившихся листьях появляются хлоротичные пятна примерно через 7 недель после прививки. Верхушечные и некоторые боковые почки становятся хлоротичными и некротическими (рис. 71, внизу справа). Симптомы менее резкие по мере того, как вирус распространяется на другие побеги. Симптомы в комплексе с другими вирусами не определялись.
Характеристика вируса. Сведения о морфологии, других свойствах вируса и серологических взаимоотношениях отсутствуют.
![Рис. 71. Симптомы кольцевой пятнистости душистой малины. Вверху слева - сорт красной малины Вашингтон; вверху справа - сорт черной малины Мунгер; внизу слева - душистая малина; внизу справа - Rubus henryi [222]](pic/000078.jpg)
Рис. 71. Симптомы кольцевой пятнистости душистой малины. Вверху слева - сорт красной малины Вашингтон; вверху справа - сорт черной малины Мунгер; внизу слева - душистая малина; внизу справа - Rubus henryi [222]
Передача. Вирус был перенесен прививкой и тремя видами тлей, которые встречаются на душистой малине: Arnphorophora parviflori Hill [128], Macrosiphurn davidsoni Mason и М. maxima Mason. Ha сеянцах черной малины симптомы обнаруживались через 7 - 10 дней после заражения тлями; на сеянцах душистой малины примерно в 2 раза позже. Эффективность заражения переносчиком была низкой; передача была более эффективной, когда в качестве тест-растений использовались сеянцы черной малины, а не душистой.
Этот вирус полуперсистентен в переносчике. Он накапливается в течение дня и сохраняется в переносчике менее одного дня. Все попытки перенести вирус механически с больных растений душистой малины на ряд травянистых тест-растений не увенчались успехом (Стейс-Смит, не опубликовано).
Распространение в естественных условиях. Сведений нет, но вирус, вероятно, распространяется тлями, которые заселяют душистую малину.
Меры борьбы. Сведений нет.
Терапия. Вирус термолабильный. Он был искоренен из растений черной малины при выдерживании их при 37° С в течение 10 дней (Стейс-Смит, не опубликовано).
Примечания. Кольцевая пятнистость душистой малины, по-видимому, ограничивается проявлением только на растениях этого вида. Вирус не передавался большой малинной тлей Amphorophora agathonica (?) Hottes, а тли, которые встречаются на душистой малине, не заселяют красную и черную малину, поэтому невероятно, чтобы это заболевание стало проблемой для промышленного возделывания малины.
Симптомы на красной малине сходны с симптомами, вызываемыми вирусом огуречной мозаики [117]. Однако невероятно, чтобы это заболевание на душистой малине вызывалось вирусом огуречной мозаики, так как попытки механической передачи были отрицательными.
Обнаружение и идентификация. По-видимому, вирус поражает только дикую душистую малину. Заметные симптомы на листьях могут служить для обнаружения и идентификации вируса.
КАРЛИКОВОСТЬ ЕЖЕВИКИ (BLACKBERRY DWARF)
Синонимы. Dwarf; Loganberry dwarf; dewberry dwarf.
История и географическое распространение. О болезни ежевики - карликовости сообщалось очень давно [83] на плантациях сортов логанова ягода, Маммот, Феноменальная в округе Сонома, Калифорния и в долине Уилламетт, Орегон. Последующими исследованиями, начатыми в 1923 г. Целлером [265, 266, 267], болезнь обнаружена в довольно ограниченных местах, где выращивались сорта Логан и Феноменальная, включая районы Калифорнии, долины Уилламетт и Ампкуа, склоны побережья Орегона и район в штате Вашингтон к западу от Каскадных гор. Последующие опыты, проведенные в штате Орегон в контролируемых условиях, показали, что болезнь вызывается вирусом, который переносится тлей [268]. Позднее Целлером и Милбратом [269] было показано, что вирус может быть перенесен прививкой на сорта Бойзен и Янг. В Калифорнии карликовость сорта Логан была описана Вильгельмом [256, 257] и Вильгельмом и др. [258 - 259а] как вирусное заболевание, переносимое прививкой, поражающее сорта Торнлесс Кори, Логан, Маммот ,и Феноменальная. Эти сорта - основные производные дикой ежевики (Rubus ursinus) Тихоокеанского побережья, естественное заражение которой было установлено в Калифорнии. Высокая восприимчивость сорта Феноменальная к карликовости в штатах Калифорнии и Орегон представляется важным фактором "истощения" и исчезновения его на промышленных плантациях ежевики.
Экономическое значение. Заметное ослабление роста и снижение продуктивности, которые были отмечены на ранних посадках сортов Логан, Маммот и Феноменальная в Калифорнии и Орегоне, приписывали карликовости ежевики [83, 256]. В середине 1920-х годов в Орегоне было установлено, что возбудителем болезни был вирус; заболевание было особенно сильным на сорте Феноменальная, но также поражало сорта Торнлесс Кори, Киттитани и Логан [267, 268]. Распространение было быстрым на сорте Феноменальная: все растения на промышленной плантации в Орегоне были заражены на третий год, если не проводилась выкорчевка больных кустов [268]. Это заболевание на сорте Логан было менее обычным и распространение болезни было более быстрым, если посадки примыкали к сорту Феноменальная. В Калифорнии заболевание имеет таких же хозяев, но симптомы несколько различны [256 - 259а]. Не вызывает сомнения, что болезнь была основным фактором как в постепенном исчезновении в Калифорнии промышленных плантаций сортов Торнлесс Кори, Маммот и Феноменальная, так и в уменьшении площади под сортом Логан. Сорт Торнлесс Логан постепенно стал более популярным, чем шиповатая форма, поскольку не поражался так сильно карликовостью. Сорта Бойзен, Гималайя, Нектар и Янг не сильно поражались при искусственном заражении, и не было установлено, чтобы они поражались в полевых условиях [256, 257].
Круг растений-хозяев. Было найдено, что сорта Торнлесс Кори, Киттитани, Логан, Маммот, Феноменальная и дикорастущая стелющаяся ежевика (Rubus ursinus Cham, et Schlecht.) с побережья Тихого океана заражены в естественных условиях [256, 268], в то время как сорта Бойзен, Гималайя, Нектар и Янг заражались искусственно [256].
В штате Орегон попытки передать прививкой вирус на сорт красной малины Кутберт и сорта черной малины Мунгер и Плам Фармер были неубедительными [268]; предварительные опыты в Калифорнии показали, что малина может быть бессимптомным носителем [258].
Симптомы. В Орегоне [268] на взрослых растениях в полевых условиях начальные симптомы карликовости проявлялись в веретеновидности и извилистости побегов со светлоокрашенными листочками, которые мельче нормальных и скорее обратнояйцевидные, чем яйцевидные. Симптомы карликовости на второй год заражения становятся более заметными вследствие карликовых побегов с укороченными междоузлиями и листьями, собранными в розетку. На этих пораженных побегах образуется по 2 - 3 почки на узел по сравнению с одной почкой на здоровых побегах. Растения, зараженные год или больше, непродуктивны. При использовании пораженных карликовостью растений для отводков побеги образуют мало или совсем не образуют корней, а верхушки, развившие корни, редко прорастают в побеги.
В Калифорнии пораженные карликовостью растения имеют слабые, веретеновидные побеги, несущие желтоватые листья, чашевидно загнутые вниз и преждевременно краснеющие и опадающие. Как и в Орегоне, в Калифорнии растения также становятся непродуктивными в течение одного года и постепенно отмирают [256].
Во время опытов по передаче вируса тлями на молодые растения сорта Логан, находящиеся под изоляторами в поле, Целлер [268] описал некроз на листьях, на которых питались вирофорные насекомые. Новые листья, развившиеся позднее, были крапчатые и морщинистые.
На этих растениях впоследствии развились симптомы карликовости, характерные при заражении в полевых условиях. Но это исследование не может рассматриваться критически, так как оно было проведено в поле (см. рис. 72).
Характеристика вируса. Сведения о морфологических особенностях, физических свойствах и серологических взаимоотношениях отсутствуют.
Передача. Передача происходит через прививку [256, 268] и, возможно, с помощью тли Chaetosiphon tetrarhodus (Walk.) (Capi-tophorus) на сорт Логан в контролируемых условиях [268]. Попытки .инфицировать здоровые растения отжатым соком из мацерированных больных листьев оказались безуспешными. В Калифорнии опыты по передаче заболевания с помощью тлей и цикадок дали отрицательные результаты (Д. Дженсен; не опубликовано).
Распространение в естественных условиях. Естественное распространение вируса более вероятно на плантациях сорта Феноменальная, чем сорта Логан [268]. В некоторых случаях, когда не было выкорчевки больных растений с плантаций сорта Феноменальная, через три года все 100% растений были заражены. Естественный переносчик не известен, но тля Chaetosiphon tetrarhodus, обитающая на розе, отмечена в качестве переносчика вируса в условиях изоляции в Орегоне [268].
Меры борьбы. Рекомендуется немедленное удаление больных растений. Восприимчивые сорта не следует высаживать вблизи дикой стелющейся ежевики Тихоокеанского побережья. Нужно высаживать здоровый посадочный материал, полученный от проверенных маточных растений из районов, где это заболевание не встречается.
Терапия. Нет сведений.
Примечания. Представляющая исторический интерес информация [S3, 256, 268] относительно широкой встречаемости карликовости в штатах Тихоокеанского побережья привела к изучению круга растений-хозяев, а сходство симптомов оправдывает включение карликовости логановой ягоды [256] как возможный синоним карликовости ежевики [69]. Необходимы дальнейшие исследования для того, чтобы установить, что эти заболевания идентичны, и определить взаимосвязь карликовости ежевики с другими вирусными болезнями ягодных культур. Сходство симптомов, образуемых на логановой ягоде карликовостью ежевики [269] и карликовостью растений Rubus, переносимой цикадкой [90, 190], вероятно, является только случайностью. Последними независимо друг от друга проведенными исследованиями (С. Вильгельм, Н. Фрезьер и X. Уильяме; не опубликовано) не установлена вирусная природа болезни Olallie [183, 184] или ее взаимосвязь с карликовостью логановой ягоды в Калифорнии.
![Рис. 72. Вверху - карликовость ежевики на плодоносящих побегах логановой ягоды: слева - задержанные в развитии побеги со слабыми плодовыми ветками и загнутыми вниз чашевидными листьями; справа - здоровый побег. Внизу - начальные симптомы заболевания на высаженной в поле логановой ягоде - веретеновидные побеги с мелкими листьями в сравнении с крупными нормальными листьями [259а, 268]](pic/000079.jpg)
Рис. 72. Вверху - карликовость ежевики на плодоносящих побегах логановой ягоды: слева - задержанные в развитии побеги со слабыми плодовыми ветками и загнутыми вниз чашевидными листьями; справа - здоровый побег. Внизу - начальные симптомы заболевания на высаженной в поле логановой ягоде - веретеновидные побеги с мелкими листьями в сравнении с крупными нормальными листьями [259а, 268]
Обнаружение и идентификация. Симптомы карликовости могут быть легко обнаружены на сильно поражаемых сортах, таких, как Торнлесс Кори, Логан, Маммот и Феноменальная. Идентичность заболевания лучше всего определяется передачей прививкой на здоровые растения сортов Логан или Феноменальная.
КАРЛИКОВОСТЬ РАСТЕНИЙ RUBUS. (RUBUS STUNT)
Синонимы. (Сокращенно RStuV.) Witches' broom, Heksenbezem, dwergziekt, Rubus Stauche, Verzwergunskrankheit, ved' miny metly, dvergsjuke.
История и географическое распространение. Хотя описание этой болезни очень краткое и недетальное, первое упоминание о карликовости растений Rubus, вероятно, было сделано де Фризом [251]. Около Бреда в Нидерландах, где расположен самый старый и основной район возделывания малины, Ритзема [201] наблюдал это заболевание в 1920 г. Вормалд и Харрис [264] сообщали об эпидемическом характере заболевания на культивируемых сортах ежевики в Англии, а Прентис [190] упоминал о сильной вспышке на гибридах малины с ежевикой, т. е. сортах Феноменальная и Логан, в той же самой стране. Согласно В. Л. Рыжкову [203а], карликовость растений Rubus была описана О. Н. Вертоградовой [250] в качестве серьезного и широко распространенного заболевания малины в Советском Союзе.
С 1920 г. Ритзема часто отмечал заболевание малины около Бреда, пока болезнь малины "ведьмины метлы" не стала бедствием примерно в 1949 г. [201]. С тех пор это заболевание стало значительно распространяться [93], а несколько лет спустя на многих молодых плантациях, которые были посажены здоровым посадочным материалом, можно было находить от 60 до 90% зараженных растений [171]. Позднее о небольших эпидемиях сообщалось из Болгарии [246] и ГДР [199]. О случайном появлении карликовости Rubus упоминалось в нескольких других европейских странах, например ФРГ, Чехословакии и Норвегии [193]. В настоящее время карликовость Rubus является серьезным заболеванием малины в западной части Польши (В. Басак, персональное сообщение). Не сообщалось о заболевании малины в западном полушарии.
Экономическое значение. Карликовость имеет большое экономическое значение в районах возделывания малины в Европе, так как потери урожая могут быть очень серьезными, когда заболевание становится массовым.
Круг растений-хозяев. а) Естественные хозяева. Вирус обнаруживается на всех основных европейских сортах малины, а также на многих видах дикой ежевики. По-видимому, все сорта малины восприимчивы, и имеется небольшое различие в чувствительности, за исключением сорта Моллинг Промис, который до некоторой степени толерантен.

Рис. 73. Карликовость Rubus на естественно зараженном растении Rubus laciniatus
б) Экспериментальные хозяева. Вирус может передаваться на Fragaria vesca и несколько промышленных сортов.
Симптомы. а) На всех видах и сортах Rubus симптомы в основном те же, т. е. множество небольших тонких побегов (рис. 73) и чрезмерное боковое ветвление всего растения вместе с филлодией и пролиферацией цветков (рис. 74, вверху).
На сорте Моллинг Промис филлодия очень редка. На плодоносящих плантациях многие больные растения, находящиеся в шоковой стадии заражения, отмирают, так как они заглушаются здоровыми. Однако растения малины, выросшие из больных корневых черенков и высаженные на правильном расстоянии одно от другого, редко отмирают и, напротив, могут проявить некоторую степень регенерации. Среди большого количества сформированных побегов некоторые становятся крупнее других и несут нормальные, но мелкие ягоды, которые трудно собирать. Плодоносящие боковые побеги на таких регенерированных растениях всегда гораздо короче, чем на здоровых. На Fragaria vesca и промышленных сортах земляники вирус вызывает "ведьмины метлы", филлодию цветков и сильное уменьшение роста (рис. 74, внизу). Зараженные растения земляники всегда отмирают в течение двух или трех лет.
б) Растения, которые уже сильно поражены резкими мозаичными болезнями, например мозаикой малины и крапчатостью листьев, значительно более чувствительны к вирусу карликовости Rubus и часто отмирают в течение нескольких лет после заражения.
Характеристика вируса. О свойствах вируса in vitro ничего не известно.

Рис. 74. Вверху - уродливые цветки и побеги красной малины сорта Редбауд, зараженного карликовостью Rubus; вверху справа показаны нормальные цветки и ягоды. Внизу - уменьшение роста растений земляники сорта Клаймакс, вызванное вирусом карликовости Rubus. Здоровое растение земляники слева
Передача. Между видами Rubus вирус переносится прививкой и цикадкой Macropsis fuscula Zett. Хотя М. fuscula не может развиваться на землянике, в лабораторном опыте она могла питаться на этом хозяине достаточно долго, чтобы передать вирус, при условии, однако, что насекомые выдерживались на инфицированных растениях малины до окончания латентного периода вируса в переносчике.
В зависимости от времени вегетационного периода, когда растения инфицируются, инкубационный период варьирует от 4 до 11 месяцев [171]. Нет различия в инкубационном периоде между растениями, которые заражаются посредством прививки, и растениями, инфицированными цикадками, так же как нет разницы в инкубационном периоде в растениях малины и земляники. Точный пороговый период приобретения вируса переносчиком не определялся. Молодые нимфы в возрасте 4 - 7 дней, собранные с больных растений, на которых они вылупились и впоследствии были перенесены на здоровые растения, не переносили вирус по мере того, как они становились взрослыми. Еще не известно, препятствовала ли получению вируса краткость периода питания или же нимфы первого возраста были не в состоянии воспринять его. Нимфы третьего и четвертого возраста, собранные с больных растений, на которых они вылупились, передавали вирус 3 - 4 недели после того, как они становились взрослыми. Таким образом, латентный период вируса может быть определен примерно равным 8 неделям. В Нидерландах молодые нимфы М. fuscula вылупляются из яиц в середине мая. Первые взрослые насекомые появляются в начале июля. Однако взрослые насекомые не способны переносить вирус до первой недели августа. Это было установлено как в полевом, так и в лабораторном опытах [91, 92]. Вирус присутствует в переносчиках до тех пор, пока они живы. Передача вируса через яйца не обнаружена.
Распространение в естественных условиях. Яйца цикадки M. fuscula перезимовывают под корой побегов растений Rubus. При большом числе яиц в искусственных культурах их легко заметить, но очень трудно найти на побегах, зараженных в естественных условиях. Поэтому трудно предотвратить распространение переносчика с посадочным материалом. Об этой цикадке сообщалось из нескольких европейских стран и Канады [245]. На культивируемых видах малины в Нидерландах цикадка встречается только в окрестности Бреда, но не в других районах возделывания малины. Однако на дикой ежевике эта цикадка встречается по всей стране, и вирус карликовости Rubus был найден на стелющихся побегах во многих местах. Эти наблюдения приводят к заключению, что зараженная ежевика не играет важной роли в сильных вспышках этой болезни вблизи Бреда. Однако в лабораторных опытах М. fuscula, взятая с нескольких видов дикой ежевики, хорошо заселяла растения малины. На Rubus caesius L. (вид дикой ежевики), обычно растущей на глинистых почвах, были найдены другие виды Macropsis, которые встречаются только на этом виде Rubus и были описаны Вагнером [252] как М. brabantica.
Вероятно, эти виды также могут переносить карликовость Rubus, так как сильные вспышки на Rubus caesius наблюдались в нескольких местах. Однако Macropsis brabantica не может развиваться на малине. Третий вид М. scotti Edw. является обычным на сорте ежевики Гималайя (R. procerus P. J. Muell.) вблизи Гоеза в юго-западной части Нидерландов. Macropsis scotti никогда не находили на малине, высаженной по соседству с сортом Гималайя, и ее неспособность развиваться на малине подтверждена лабораторными опытами. Согласно Леггу, как указывал Кадмен [45], М. scotti также может переносить вирус карликовости Rubus. Однако в Нидерландах вирус никогда не был найден на ежевике сорта Гималайя.

Рис. 75. Первые симптомы карликовости Rubus на красной малине. Многочисленные слабые побеги развиваются из корневых почек; предшественник 'ведьминой метлы'
На основании полевых наблюдений установлено, что заражение главным образом происходит на плантациях малины в августе и сентябре первого вегетационного периода. Первые симптомы (многочисленные слабые побеги, развивающиеся из корневых почек) появляются в августе и сентябре следующего года (см. рис. 73 и 75). На третий год после посадки летом появляются типичные симптомы "ведьмины метлы": короткие плодовые боковые веточки с ненормальными цветками и множеством слабых и тонких побегов. Во время одной из вспышек заболевания вблизи Бреда можно было наблюдать, что симптомы никогда не развивались в первом вегетационном периоде, если использовался здоровый посадочный материал. Поэтому, если больные растения обнаруживаются в первый вегетационный период, можно быть уверенным, что они высаживались уже зараженными. Во время обследования в 1953 и 1954 гг. было установлено, что растения на старых плантациях были значительно более устойчивыми, чем на молодых плантациях [172]. На ряде ферм были обнаружены 8 - 10-летние посадки, содержащие примерно 10% больных растений, тогда как на этих же фермах двухлетние посадки, расположенные рядом со старыми, имели от 50 до 80% больных растений.
Это может быть объяснено тем, что вновь посаженные растения малины начали расти позднее, чем старые растения, и, таким образом, являются физиологически более молодыми к августу и сентябрю, т. е. к тому времени, когда цикадки способны переносить вирус. Молодые побеги на таких посадках сохраняют листья очень долго, тогда как молодые побеги на старых плантациях начинают терять листья в июле. Наблюдения действительно показали, что популяция цикадок на старых здоровых растениях уменьшалась очень быстро после конца июля, в то время как на молодых растениях, которые были свободны от цикадок при посадке, Macropsis fuscula можно было находить до октября. Больные растения на старых плантациях являются хорошими растениями-хозяевами для цикадок, так как их многочисленные побеги продолжают рост до поздней осени. Однако, по-видимому, они имеют небольшое значение для распространения инфекции на таких старых плантациях.
Меры борьбы. Яйца цикадки М. fuscula могут быть легко убиты опрыскиванием минеральными маслами зимой, тогда как с нимфами можно бороться с помощью опрыскивания паратионом* в мае.
*(В СССР не применяется. - Ред. )
В Нидерландах Консультативный Совет провел кампанию, в которой каждый фермер был обязан провести опрыскивание своей плантации малины минеральными маслами зимой и паратионом* в мае [172, 201, 211, 212].
*(В СССР не применяется. - Ред.)
Садоводам рекомендовалось также удалять и уничтожать все больные растения. Эта кампания дала хорошие результаты в борьбе с переносчиком вируса [174].
Терапия. От карликовости Rubus растения можно легко вылечить с помощью тепловой обработки [244]. Например, окорененные зараженные побеги малины были освобождены от вируса карликовости Rubus после прогревания в воде с температурой 46°С в течение 2 - 3 ч.
Примечания. Хотя землянику выращивают очень часто вблизи Бреда, заражение карликовостью Rubus никогда не наблюдалось на этой культуре.
Длительный период инкубации и симптомы на цветках земляники, зараженной в искусственных условиях, подтверждают, что карликовость Rubus и позеленение лепестков земляники [188] вызываются различными вирусами.
По мнению авторов, карликовость Rubus является единственным известным заболеванием, имеющим важное экономическое значение для рода Rubus. Было доказано, что оно вызывается вирусом, переносимым цикадкой. Однако болезнь Пирса, вызываемая другим вирусом, переносимым цикадкой, по-видимому, встречается в латентной форме на растениях Rubus vitifolius Cham, et Schleht [98]. Симптомы "ведьмины метлы" на черной малине наблюдались в США Цунделем [270] в Пенсильвании и Конверсом (персональное сообщение) в Мичигане в 1963 г. В последнем случае пораженные растения настолько быстро погибали, что передача через прививку не могла быть проведена. Никольс и др. [183] сообщили о пролиферации побегов у ежевики сорта Олалли в Калифорнии.
Симптомы напоминали симптомы, вызываемые некоторыми переносимыми цикадками вирусами, но последние недавно проведенные исследования не смогли установить связь этого заболевания с вирусом.
Обнаружение и идентификация. Обнаружение по симптомам: образование многочисленных побегов вместе с ветвлением боковых веточек. Филлодия - наиболее типичный симптом. Некоторые сорта малины и ежевики могут регенерировать в высокой степени. Такие регенерированные растения не проявляют заметной деформации цветков и поэтому должны проверяться прививкой на растения чувствительных сортов малины, таких, как Радбауд и Моллинг Ландмарк.
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://wine.historic.ru/ 'Виноделие как искусство'