Основные принципы культивирования дрожжей
Специфические особенности производства шампанского в непрерывном потоке потребовали принципиального изменения способов культивирования дрожжей с целью повышения их функциональной деятельности.
Широкое внедрение непрерывных процессов на различных стадиях производства Советского шампанского обусловило необходимость изучения основных закономерностей развития дрожжевых организмов с целью возможного изменения активности жизненных процессов и их направленности.
В условиях шампанского производства дрожжи проявляют активность "на границе своих биологических возможностей". Шампанские расы дрожжей должны быть хорошо приспособлены к "жестким" условиям среды обитания.
Жизнедеятельность дрожжей определяется степенью их приспособленности к внешним условиям. Значительную роль в производстве шампанского играют состав среды и условия культивирования дрожжей. Существенное влияние при этом оказывает температурный режим, содержание кислорода, а также наличие ростовых веществ в производственном субстрате.
Для осуществления дрожжами энергетического обмена питательная среда должна быть полноценна по своему составу. В процессе жизнедеятельности дрожжи непрерывно подвергаются разнообразным превращениям. При этом происходит энергичный рост клетки, увеличение ее массы, формирование клеточной структуры и, наконец, размножение. Истинные дрожжи семейства сахаромицетов размножаются почкованием. В процессе размножения ядро делится на два, одно из них вместе с частью цитоплазмы и другими клеточными элементами переходит в молодую, вновь образующуюся дочернюю клетку. При почковании клетки происходит также деление митохондрий, имеющих вид либо небольших гранул, палочек или нитей, либо цепочек из отдельных зерен. Митохондрии распадаются на отдельные части, причем в каждую клетку (дочернюю и материнскую) обязательно попадает одинаковая часть отделившихся митохондрий. Почкующиеся материнские и дочерние клетки обладают гомогенной плазмой и тонкой оболочкой. Эти молодые клетки содержат большой запас резервных углеводов и мелкие вакуоли.
Для нормального роста, развития и размножения дрожжам необходима энергия, которую они получают в процессе дыхания и брожения. Сбраживание углеводов дрожжами при шампанизации - процесс анаэробный. Шампанские расы дрожжей - организмы в основном факультативно анаэробные. В анаэробных условиях брожение идет интенсивно, однако рост и размножение их заторможены. При этом процессы синтеза в дрожжевой клетке протекают значительно медленнее, чем при дыхании, так как на 1 моль потребленной глюкозы образуется лишь 2 моля аденазинтрифосфата (АТФ), тогда как при дыхании - 38 молей. Таким образом, брожение является значительно менее совершенным способом получения энергии, чем дыхание, и сопровождается большой тратой питательных веществ [61].
При наличии в среде кислорода активность брожения значительно ослабевает, а дыхание возрастает. Повышение активности дыхания способствует ускорению роста и размножения дрожжей.
Начало изучения влияния кислорода на спиртовое брожение было положено Л. Пастером в 1857 г., который впервые установил явление, позднее названное Варбургом "эффектом Пастера". Оно выражается прежде всего в угнетении анаэробного брожения дыханием, в значительном снижении потребления субстрата - глюкозы и уменьшении образования этанола. Это явление послужило в дальнейшем основой для развития учения об эффекте Пастера как об одном из важнейших регуляторных механизмов энергетического обмена дрожжевой клетки.
Механизм действия кислорода на дрожжи очень сложен. Он охватывает все функции клеток. При изучении процесса дыхания было установлено, что дрожжи обладают характерным спектром цитохромов [158]. При изменении условий культивирования (аэрации) отмечены большие колебания системы ферментов дыхания дрожжей. В процессе аэробиоза в клетках происходит образование митохондрий, увеличение активности ферментов электроннотранспортной цепи, цикла Крепса, а также усиление синтеза АТФ [165].
Работами по изучению роста и размножения дрожжей в условиях периодического культивирования [119, 110, 106] установлено, что дрожжи при этом очень короткое время находятся в фазе логарифмического роста и нередко переходят в угнетенное состояние. Кроме того, дрожжи каждой из последующих генераций по своим свойствам, как правило, отличаются от предыдущих. В результате этого получаемая масса дрожжей не является однородной. Клетки зрелых дрожжей отдельных разводок часто обладают различной физиологической активностью и не одинаково устойчивы к неблагоприятным воздействиям производственных факторов. Более того, длительное культивирование дрожжей при периодическом способе их воспроизводства при одной и той же необновляемой среде, обогащенной продуктами метаболизма, приводит в итоге к угнетению культуры и значительному ослаблению ее биологической активности. Дрожжи, приготовленные периодическим способом, физиологически недостаточно активны, плохо адаптированы к специфическим жестким условиям производства шампанского. При внесении в резервуары они нередко вызывают вялый процесс вторичного брожения, что создает предпосылки к развитию нежелательной посторонней микрофлоры, в частности диких дрожжей, молочнокислых бактерий и др. Таким образом, существующие до настоящего времени периодические способы производства чистых культур дрожжей в винодельческой промышленности не удовлетворяют возросшим требованиям, выдвигаемым в последнее время в связи с массовым внедрением в промышленность нового, прогрессивного метода шампанизации вина в непрерывном потоке.
В многочисленных работах по изучению поведения дрожжей при их культивировании, проведенных как в лабораторных, так и в производственных условиях, исследователи обычно суммарно учитывают процессы размножения и брожения всей популяции дрожжей в той или иной среде. В то же время для полноценного управления бродильными процессами, особенно при переводе их на непрерывные, несомненный интерес представляет более углубленное понимание поведения отдельных дрожжевых клеток и корреляции между процессами их размножения и брожения при жизнедеятельности культуры в производственных субстратах.
Установлены закономерности размножения и брожения дрожжей в виноградном сусле и бродильной смеси [50].
Подсчет клеток в субстратах с помощью счетной камеры производился через возможно более короткие отрезки времени (обычно через 3 ч) и, как правило, сопровождался подсчетом почек (пробы фиксировались серной кислотой, разведенной водой в соотношении 1:5). Полученные данные обрабатывались по следующей формуле:
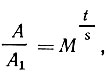
где A1 - начальное число клеток в единицу времени;
А - число клеток в момент времени t;
s - продолжительность образования и полного оформления дочерней клетки (скорость процесса почкования);
M - энергия размножения популяции;
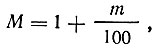
(m - процентное число почкующихся клеток).
Подставляя в формулу экспериментально найденные величины, вычисляли путем логарифмирования скорость почкования s.
Сопоставление составленных по экспериментальным данным кривой М с кривой размножения дрожжей дает возможность более точно судить, за счет чего идет нарастание клеточного материала: большего ли вовлечения числа клеток в процесс почкования или большей скорости формирования почек в клетке. Это представляет собой интерес при изучении влияния различных факторов на процесс размножения дрожжей.
Одновременно в модифицированных бродильных приборах по объему выделенной углекислоты вычисляли количество сахара (в мг), сброженного дрожжевой популяцией в 100 мл среды. Затем вычисляли энергию брожения одной дрожжевой клетки в каждый момент наблюдения и выражали ее в миллионных долях микрограмма сахара (у), сброженного в течение часа.
Исследования показали, что у дрожжей расы Штейнберг при высеве в сусло максимум брожения одной клетки наступает при наличии в среде 75% клеток в процессе почкования, при почковании около 2 ч сбраживается около 21γ сахара.
Максимум брожения одной дрожжевой клетки наступает при стационарном процессе ранее максимума брожения всей популяции, т. е. главного брожения. В момент наивысшей активности брожения в сусле клетка винных дрожжей в 1 ч выделяет количество углекислоты, во много раз (10-20) превышающее объем ее тела.
При проведении производственного микробиологического контроля определение процента почкующихся клеток в отсутствие нарастания процента мертвых клеток (окраска метиленовой синью в концентрации 1:10000) свыше принятых норм является достаточно надежным критерием активности культуры.
Наибольшая активность брожения клетки наступает раньше накопления ею максимального количества гликогена. Указанное положение находится в противоречии с установившимся и вошедшим в руководства взглядом, согласно которому реакция на гликоген берется как основная при оценке бродильной активности заводских дрожжей ряда производств. Роль гликогена в дрожжевых клетках еще до конца не раскрыта. Если присоединиться к точке зрения некоторых авторов [174], которые рассматривали образование гликогена как первую фазу брожения, то отсутствие его в больших количествах в клетке в период наиболее энергичного брожения становится понятным. Растворимый сахар среды диффундирует внутрь дрожжевой клетки и в ней превращается в гликоген, клетка, находясь в состоянии высокой активности, быстро расщепляет его и фосфорилирует продукты гликогенолиза; гликоген не успевает накопиться в больших количествах. Когда же активность зимазы начинает уменьшаться, гликоген откладывается в качестве резервного продукта. Очевидно, оценка максимальной активности дрожжей по наибольшему количеству гликогена не является достаточно точной.
Необходимо подчеркнуть, что активность дрожжевой клетки в большей степени зависит от ее ориентации в среде. Клетки, находящиеся во взвешенном состоянии, почкуются более энергично и образуют больше алкоголя, чем находящиеся в осадке. Различие это, незначительное в начальной стадии брожения, со временем усиливается во много раз.
При шампанизации вина присутствующие в нем компоненты, особенно спирт, оказывают возрастающее депрессирующее влияние на размножение дрожжей. Это наиболее резко проявляется в падении скорости почкования клеток. Количество же клеток, принимающих участие в размножении (процент почкующихся клеток), уменьшается менее значительно. Происходит также ускорение процесса отмирания клеток.
Таким образом, по существу дрожжи в резервуарах проявляют свою активность на пределе биологических возможностей. Достаточно незначительного ухудшения условий, и дрожжи окажутся бессильными выбродить сахар бродильной смеси до нормы.
При периодическом культивировании дрожжей размножение клеток, зависящее от ряда факторов, протекает в основном в три фазы: начальную стационарную (лаг-фазу), логарифмическую и фазу отрицательного ускорения (гибели клеток). Когда культуру микроорганизма переносят в новые условия среды, клетки, как правило, не сразу начинают размножаться, а некоторое время находятся в стадии задержки роста. Этот период носит название лаг-фазы. Происходит как бы приспособление клеток к новым условиям среды. Изучение сущности процессов, происходящих при лаг-фазе, представляет значительный теоретический и практический интерес и является предметом продолжающихся исследований.
В настоящее время установлено, что в среде с недостаточно полноценными питательными свойствами лаг-фаза удлиняется. Физико-химические условия также оказывают на лаг-фазу большое влияние. В частности, аэрация среды при культивировании аэробных микроорганизмов укорачивает лаг-фазу. При культивировании анаэробных микроорганизмов аэрация и высокий окислительно-восстановительный потенциал удлиняют лаг-фазу. Значительное влияние оказывает и температура среды. Лаг-фаза существует и при наиболее оптимальных условиях культивирования. Более того, при посеве молодой активной культуры, находящейся в фазе логарифмического роста, лаг-фаза все же наблюдается. Следовательно, она бывает различной на одной и той же среде и обусловливается не только внешними, но и внутренними причинами. Клетки при этом находятся в активном взаимодействии со средой. В этот период анаэробные микроорганизмы выделяют в среду мощные восстановители, снижающие окислительно-восстановительный потенциал до оптимальных значений для данного вида.
Потом наступает фаза логарифмического (экспоненциального) роста, когда число клеток увеличивается в геометрической прогрессии.
Переход логарифмической фазы в стадию задержки роста связан или с накоплением продуктов метаболизма, или с недостаточным количеством элементов питательной среды. Созданием соответствующих условий культивирования логарифмическую фазу роста можно продлить. По мере истощения запасов питательных веществ и накопления продуктов метаболизма размножение клеток замедляется, а затем и прекращается. Вегетативные клетки при этом либо отмирают, либо переходят в покоящиеся формы. Для предупреждения процесса преждевременного старения курьтуры ее необходимо длительное время поддерживать в стадии экспоненциального роста. Практическое осуществление этого возможно лишь при использовании проточной, непрерывно обновляемой среды [46].
Важными факторами, вызывающими повышение активности размножения и брожения дрожжей, являются непрерывное перемешивание, а также наличие растворенного кислорода в культуральной жидкости. Аэрация, совмещенная с перемешиванием, способствует большему усвоению дрожжами кислорода и, следовательно, значительному накоплению их биомассы [160]. При этом существенное влияние на скорость размножения дрожжей оказывает величина окислительно-восстановительного потенциала, который представляет собой "состояние динамического равновесия культуры между активностью энзимов клетки и веществами среды".
Исследования по приготовлению дрожжевой разводки непрерывным способом показали преимущество его перед периодическим, заключающееся в эффективности естественного отбора клеток, наиболее приспособленных к специфическим условиям производства, и изменении их свойств в нужном направлении [119].
Изучение процессов непрерывной шампанизации вина показало, что качество, количество и способность дрожжей к воспроизводству, а также поддержание постоянства их концентрации в бродильной батарее являются важнейшими факторами, определяющими весь ход технологического процесса [21]. В условиях непрерывного культивирования по сравнению с периодическим при одинаковых температурах процессы протекают быстрее.
Теоретическое и практическое обоснование непрерывного культивирования микроорганизмов дано в работах многих авторов. Подтвержден основной вывод о возможности размножения бактерий, бацилл и дрожжей непрерывным методом [168]. Установленные закономерности размножения дрожжей, их адаптации к условиям непрерывного процесса послужили основой для организации производственного получения кормовых дрожжей.
Из теоретических исследований в области непрерывного культивирования микроорганизмов представляют интерес данные, свидетельствующие о том, что при благоприятных условиях проточная культура обладает способностью саморегуляции. Количество микробных клеток при этом можно поддерживать на любом уровне, обеспечив соответствующую скорость поступления питательных веществ [169, 170].
При изучении влияния внешней среды на размножение дрожжей в условиях периодического культивирования установлено, что процесс роста и размножения клеток происходит в среде, непрерывно изменяющейся по составу. Это приводит в конечном счете к нарушению обмена веществ и ослаблению их физиологической активности [106]. Характерной особенностью дрожжей при непрерывном культивировании является интенсификация процесса: размножение дрожжей при одинаковом исходном составе питательной среды и одинаковой температуре происходит в 20 раз быстрее, чем при периодическом [162].
Таким образом, культивирование микроорганизмов в проточных, непрерывно обновляемых средах является одним из наиболее эффективных средств управления обменом веществ, ростом и их развитием [167, 46].
Следует отметить, что понятия "проточное" и "непрерывное" культивирование не всегда совпадают. В отдельных случаях можно добиться непрерывности процесса без применения проточных сред, например при производстве дрожжей" отъемным способом. После того как концентрация дрожжей в субстрате достигнет определенного уровня, 2/3 объема разводки отбирают и ферментер вновь дополняют свежей питательной средой. Следовательно, среда поступает периодически, а размножение происходит непрерывно. Концентрация клеток при периодических отъемах и доливках колеблется в широких пределах. Если приток питательной среды и отъем разводки производить непрерывно, то периодический способ станет типично гомогенно-непрерывным. Размножение микроорганизмов этим способом осуществляется в одной емкости при тщательном перемешивании среды, обеспечивающем гомогенность культуральной жидкости и постоянство концентрации биомассы.
Градиентно-непрерывный способ основан на принципе культивирования микроорганизмов в батарее последовательно соединенных ферментеров. Сущность этого способа заключается в том, что в отдельных ферментерах создаются определенные градиенты в составе питательной среды и физиологическом состоянии клеток по направлению движения потока.
Для теоретического обоснования непрерывных процессов выведены некоторые математические закономерности роста культуры.
При гомогенно-проточном культивировании непрерывность процесса обеспечивается динамическим равновесием между приростом биомассы и коэффициентом разбавления
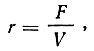
где F - поступление в ферментер питательной среды, мл/ч;
V - рабочий объем ферментера, мл.
Этот коэффициент характеризует сменяемость объема культуральной жидкости в течение часа.
Скорость роста биомассы в единицу времени проточной культуры выражают уравнением
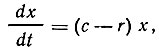
где c - коэффициент скорости роста культуры;
r - коэффициент разбавления;
x - биомасса, или количество клеток в единице объема культуральной жидкости.
При установившемся потоке количество клеток в единицу времени является величиной постоянной 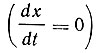 до тех пор, пока c - r = 0. Такое равенство может быть достигнуто различными путями.
до тех пор, пока c - r = 0. Такое равенство может быть достигнуто различными путями.
При гомогенно-проточном культивировании обычно используют установки типа турбидостат и хемостат. Принцип регулирования численности популяции в аппаратах типа хемостат основан на том, что прирост биомассы пропорционален количеству потребленного вещества:
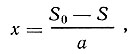
где x - биомасса;
S0 и S - исходная и остаточная концентрация питательных веществ:
a - количество питательных веществ субстрата, израсходованных на образование единицы микробной биомассы.
Для хемостата применяются среды с большим избытком питательных веществ, за исключением одного, лимитирующего рост. Следовательно, удельная скорость роста является функцией лимитирующего вещества. Концентрация его (сахар, витамин, аминокислота и пр.) в зависимости от требуемых условий может быть изменена в широких пределах. Если скорость притока среды не очень велика, это лимитирующее вещество потребляется практически полностью. По мере увеличения притока и, следовательно, коэффициента разбавления возрастает и количество вещества, которое культура успевает перерабатывать. В связи с этим количество клеток в культуральной жидкости снижается и может дойти до нуля. Когда коэффициент разбавления превысит некоторую критическую величину, культура практически будет вымыта из аппарата. В этом случае S0 и S будут равны между собой. Однако снижение численности клеток вызовет увеличение энергии их размножения до тех пор, пока она не уравняется со скоростью разбавления. После установления равновесия между энергией размножения и скоростью разбавления количество клеток в культуральной жидкости устанавливается на определенном уровне. В случае замедления притока количество клеток возрастет, и этот уровень повысится.
Таким образом, между численностью микробных клеток и скоростью их размножения существует обратная зависимость. А численность клеток в свою очередь определяется в основном скоростью притока среды.
В аппаратах типа турбидостат микроорганизмы благодаря использованию богатых питательных сред размножаются с максимальной скоростью. Количество биомассы культуры поддерживают на заданном уровне путем обеспечения необходимого притока питательной среды. В отличие от хемостата здесь отсутствуют лимитирующие факторы.
Принцип хемостата широко применяют в научно-исследовательской работе и производственной практике, а принцип турбидостата имеет ограниченное распространение.
Непрерывное культивирование микроорганизмов осуществляется в одну или две фазы. При однофазном процессе превалирует размножение и накопление биомассы. Культивирование в две фазы проводят в случае необходимости получения в основном конечных продуктов жизнедеятельности микробной клетки.
Для проведения однофазного процесса обычно применяют гомогенно-непрерывный способ культивирования, а для многофазного процесса - градиентно-непрерывный способ. Закономерности роста культуры при этом более сложны, чем при гомогенно-проточном, так как в этих условиях питательная среда во второй и последующие ферментеры поступает с некоторым содержанием микробных клеток. Если первый ферментер батареи условно считать гомогенно-проточным, то скорость роста биомассы в единицу времени выражается уравнением 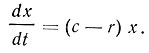 . Во втором и последующих ферментерах увеличение количества клеток может быть представлено следующей зависимостью:
. Во втором и последующих ферментерах увеличение количества клеток может быть представлено следующей зависимостью:
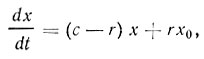
где x0 - биомасса, или количество клеток, в единице объема культуралыюй жидкости, поступающей из предыдущего ферментера.
Численность клеток остается постоянной 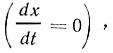 , если
, если 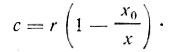 . В данном случае соотношение между c и r может изменяться в широких пределах. С уменьшением коэффициента разбавления возрастает концентрация клеток в субстрате и наоборот.
. В данном случае соотношение между c и r может изменяться в широких пределах. С уменьшением коэффициента разбавления возрастает концентрация клеток в субстрате и наоборот.
Как известно, эффективность различных систем аппаратуры и способов культивирования характеризуется показателем θ, который является величиной, обратной коэффициенту разбавления, т. е. 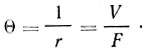 . Величина θ показывает оборачиваемость ферментера. При батарейной установке продуктивность культуры P может быть выражена следующим уравнением:
. Величина θ показывает оборачиваемость ферментера. При батарейной установке продуктивность культуры P может быть выражена следующим уравнением:
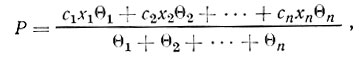
где цифровые индексы указывают порядковый номер ферментеров.
Существенно, что в последних ферментерах культура может не только не находиться в стадии размножения или брожения, но даже автолизироваться и отмирать. Следовательно, коэффициенты скорости роста в этом случае могут иметь нулевое или отрицательное значение. Таким образом, количество ферментеров следует подбирать с учетом требования производства. При этом необходимо учитывать также и физиологическую активность культуры, которая может быть выражена количеством переработанного за 1 ч субстрата (образовавшихся продуктов) в пересчете на единицу биомассы. Как правило, физиологическая активность молодых, быстро размножающихся особей бывает выше, чем старых, медленно растущих клеток. Однако при двухфазном процессе наблюдается обратная закономерность. В этом случае способность образовывать продукты обмена у молодых клеток гораздо ниже, чем у физиологических зрелых. Следовательно, протекающие при непрерывном культивировании процессы условно могут быть разделены на две стадии:
накопление биомассы, когда рост является результатом обмена веществ без видимого синтеза продуктов метаболизма;
рост биомассы с накоплением значительных количеств метаболитов микробной клетки.
Очевидно, что в условиях шампанского производства культивирование дрожжей в непрерывном потоке целесообразно проводить в направлении получения как физиологически активной биомассы, так и некоторого количества продуктов метаболизма.
До разработки и внедрения в промышленность способа шампанизации вина в непрерывном потоке чистую культуру дрожжей готовили периодическим способом. Дрожжи обычно отбирали из акратофора, имеющего лучшие показатели брожения, и вносили в дрожжанки со стерильной питательной средой. На некоторых заводах практиковали отъем дрожжей не из акратофоров, а из готовой дрожжевой разводки, находящейся в стадии бурного брожения.
Периодический способ размножения дрожжей, как показали исследования, обладает рядом существенных недостатков. Дрожжи непродолжительное время находятся в фазе логарифмического роста. В связи с тем что состав и температура культуральной жидкости изменяются в процессе культивирования, дрожжи каждой из последующих генераций, как правило, несколько отличаются от предыдущих по своим свойствам. Размножение дрожжей происходит в среде, в которой в результате их жизнедеятельности питательные свойства ухудшаются, а факторы, ограничивающие рост, усиливаются. В связи с этим получаемая масса дрожжей не однородна по своим свойствам. Клетки зрелых дрожжей отдельных разводок часто обладают различной физиологической активностью и неодинаковой устойчивостью к неблагоприятным воздействиям производственных факторов. Длительное культивирование дрожжей на одной и той же необновляемой среде, обогащенной продуктами метаболизма, приводит к угнетению культуры и значительному ослаблению ее биологической активности. При этом они плохо адаптированы к специфическим условиям производства и нередко вызывают вялый процесс вторичного брожения, что создает предпосылки к развитию инфекции, в частности молочнокислыми бактериями.
Следовательно, существовавший ранее способ производства дрожжей не удовлетворял в полной мере возросшим требованиям, связанным с внедрением в промышленность прогрессивного способа шампанизации вина в непрерывном потоке. Как отмечалось, качество и интенсивность воспроизводства дрожжей, а также поддержание постоянной концентрации их в шампанизируемом вине являются важнейшими факторами, определяющими весь ход технологического процесса. В связи с этим был разработан полунепрерывный способ культивирования дрожжей, примененный в типовой установке. Проведенное в дальнейшем усовершенствование технологии, направленное на повышение эффективности производства, а также необходимость распространения принципа непрерывности на все подготовительные и заключительные стадии основного процесса вызвали дополнительные требования к культивированию дрожжей. Так, оптимизация режима биологического обескислороживания и термической обработки купажа, уменьшение сахаристости бродильной смеси и изменение принципа подачи ее на шампанизацию, выдержка ликеров в потоке обусловили существенное повышение качества продукции, но вместе с тем значительно затруднили проведение вторичного брожения.
В этих условиях дрожжи оказались на пределе своих физиологических возможностей, что настоятельно диктовало необходимость разработки новых условий их культивирования. Для решения этой задачи был проведен анализ существующих способов непрерывного культивирования дрожжей, показавший необходимость принципиальной переработки их применительно к шампанскому производству.
Решение проблемы непрерывного культивирования дрожжей, как известно, сопряжено с рядом трудностей, обусловленных прежде всего быстрым развитием инфекции, что в условиях шампанского производства по-видимому является следствием сравнительно благоприятных условий для этого и недостаточно высокой активности применяемых дрожжей.
Разработан способ культивирования дрожжей в непрерывном потоке, который обеспечил накопление клеток в физиологически активном состоянии, адаптированных к жестким производственным условиям среды обитания, в количестве, гарантирующем нормальное проведение вторичного брожения [122].
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://wine.historic.ru/ 'Виноделие как искусство'